|
Viruslarda Mutasyonlar ve Rekombinasyonlar Prof. Dr. Mustafa Arda Ankara Üniversitesi Veteriner Fakültesi 01. Mutasyonlar 01. Mutasyonlar 01.01. Genel Bilgiler Viruslar, diğer
mikroorganizmalardan daha fazla genetik varyasyonlara açıktırlar ve maruz kalırlar. Bu
değişiklikler, genellikle, genetik yapıda oluşan ve fenotipe de etkileyen mutasyonlar sonucunda ortaya çıkmaktadırlar.
Viruslar uygun (permisif) canlı
sistemlerde (deneme hayvanları, embriyolu yumurtalar, hücre kültürleri) üreyerek
kısa bir süre içinde milyonlarca hatta milyarlarca yeni nesil oluşturabilmektedirler.
Gayet doğaldır ki, DNA veya RNA'nın bu kadar fazla replikasyonları sırasında,
genetik düzeyde kendiliğinden (doğal
veya spontan olarak) bazı değişmeler
meydana gelmektedir. Bazen de, virusların üretildikleri ortama ilave edilen bir kısım mutajenik maddeler, mutasyonlara yol açarak,
böyle değişikliklerin oranını daha da artırmaktadır. Spontan mutasyonların oranı,
normal koşullar altında, genellikle, çok sınırlı (10-6 - 10-8
arası) olmasına karşın, mutajenik maddelerin katılımı ile bu oran çok
yükselmektedir (10-3 - 10-4). Mutasyonlar sonucunda, orijinallerine göre
değişik genotipte ve buna bağlı olarak
fenotipte gözlemlenen ve saptanan yeni nesiller (mutantlar)
meydana gelirler. Ancak, mutasyonları tam veya kısmen de olsa önlemek ve düzeltmek
için, üredikleri sistemlerde, bazı mekanizmalar da yok değildir. Bu düzeltme sistemleri, virusların
genomlarından ziyade, hücrelerde bulunmaktadır. Bunlar da çoğu zaman yetirsiz
kalmaktadırlar. Çünkü, bazı virusların replikasyonları ve ekspresyonlarında
görevli enzimlerinin böyle bir aktivitesi bulunmamaktadır. Bu olgu, bakterilerde çok
daha etkindir. Virusların fazla
sıklıkta mutasyonlarla karşı karşıya gelmelerinin diğer nedenleri arasında,
bunların genetik materyallerinin daha komplike bir karakter taşımasıdır. Şöyle ki,
virusların DNA veya RNA karakterinde bir genetik materyale sahip olmaları yanı sıra,
tek veya çift iplikçikli, segmentli-segmentsiz, lineer-sirküler, pozitif veya negatif
polariteli, bazılarının süpersarmal bir özellik taşımaları, ayrıca replikasyon
yerlerinin nukleus veya sitoplasma olmaları, virus familyaları arasında replikasyon
sırasında değişik enzimlerin ardışık fonksiyonel bulunmaları ve diğer nedenler bu
mutasyonların oluşmasında kolaylaştırıcı faktör olarak etkilemektedirler. Mutasyonların
bazıları hafif veya belli-belirsiz (silent) olmakta,
genomda önemli değişiklikler yapmamakta ve tamiratı da kolayca yapılabilmektedir.
Buna karşın bir kısmı ise, genetik materyallerde önemli bozukluklar yaparak
(virulens, patojenite, antijenite ve diğer önemli faktörlerde), orijinal (parental)
viruslara oranla farklı genotipte mutantların ortaya çıkmasına ve hatta bazı
mutasyonlar da virusların ölmesine (letal
mutasyon) yol açmaktadırlar. Bu son türdeki mutasyonlar, nukleik asitlerin temel
niteliğindeki genlerinde oluşan, köklü ve tamiri yapılamayan değişikliklerden
kaynaklanmaktadır. Silent mutasyonlar,
genellikle, viral genomda, bir bazın girmesi veya çıkması ile karakterize olan
türdeki varyasyonları kapsamakta ve bir kaç generasyon sonra tamir edilebilmektedirler. Bazı değişiklikler
de, genetik defektif virusların (bunlar,
helper viruslarının yardımı ile
hücrelerde replike olabilir ve çoğalabilirler) ortaya çıkmasına yol açarlar. Temel genlerin
dışındakilerde oluşun mutasyonlar bazen, antiviral
ilaçlara karşı virusların direnç kazanmasına ve antijenite de önemli
değişikliklerin ve buna bağlı olarak ta alt
serotiplerin meydana gelmesine neden olabilmektedirler. Nukleik asitlerin
yapılarını oluşturan bazların sırasında ve türlerinde bir çok tarzda
değişiklikler görülebilmektedir. Bunların arasında başlıca, 1) Nukleik asitlerin
normal baz sıraları arasından bir baz çiftinin
çıkması (delesyon) veya baz sıraları arasına bir baz çiftinin girmesi (insersiyon)
sonucunda mutasyonlar ortaya çıkabilirler (çift iplikçikli genoma sahip viruslarda). 2) Baz sıraları
arasına bir bazın girmesi veya baz
sıralarından bir bazın çıkması
sonucunda mutasyonlar görülebilir (tek veya çift iplikçikli genomlarda). 3) Bir baz çiftinin
yerini yine aynı türden diğer bir baz çiftinin (transisyonel mutasyonlar) veya farklı türden
bir baz çiftinin alması (transversiyonal
mutasyonlar) mutasyonlara yol açabilir (çift iplikçikli genoma sahip viruslarda)
(nokta mutasyonları). 4) Aynı iplikçik
üzerinde yan yana bulunan bazlar arasında kovalent bağlar kurularak ortaya çıkan
birleşmeler (genellikle, timin dimerleri arasında
oluşan dimerizasyon gibi) mutasyonlara
neden olabilmektedir (tek veya çift iplikçikli genomlarda). 5) Tek veya çift
iplikçikli genomlarda, değişik tarzlarda (baz-şeker bağlarının kopması, bazlar
arası karşılıklı hidrojen bağlarının parçalanması, çapraz bağların
kurulması, şeker-fosfat molekülleri arasındaki bağların kopması, ve diğer
nedenler) mutasyonlar (hafif veya önemli) ortaya çıkabilmektedir. Yukarda, genel hatları
ile belirtilen mutasyonlar, virusların hücreler içindeki replikasyonları ve
ekspresyonları (transkripsiyon ve translasyon) sırasında kendiliğinden (spontan olarak) veya çeşitli mutajenik maddelerin (mutajenler: Fiziksel mutajenler, "UV-ışınları, X-ışınları" veya kimyasal mutajenler" nitröz asidi, hidroksil amin, alkilen maddeler, baz analogları, akridinler, vs") etkisi altında meydana
gelmektedirler. Mutasyonlara, RNA
viruslarında, DNA viruslarından daha fazla rastlanılmaktadır. Bunun başlıca
nedenleri arasında, RNA viruslarında, replikasyonda görev alan RNA polimeraz enziminin
(RNA'ya bağımlı RNA polimeraz) geriye yönelik
düzeltme mekanizmasının bulunmamasıdır. Bu nedenle replikasyon sırasında
oluşan hatalar (yanlış bazların sıraya girmesi ve bunların düzeltilememesi) aynen
kalırlar. Ayrıca, sellüler orijinli RNA polimeraz enziminin de yanlışı düzelterek
doğru bazı yerine koyma kapasitesi bulunmamaktadır. Bu durumlara bağlı olarak RNA
viruslarında 2500-10000 polimerize nukleotide bir hata isabet etmektedir. Ayrıca,
küçük RNA viruslarında her bir replikasyonda bir veya birden fazla yanlış
nukleotidin sıraya girebileceği belirtilmiştir. Mutasyonlar, diğer
özellikleri yanı sıra, plak formasyonlarında da
kendini belli eder ve değişik morfolojiye sahip plaklar meydana gelebilirler. Böylece heterojen virus populasyonlarının
oluşmalarına yol açarlar. Bu durum da, genellikle antijenitede oluşan değişiklikleri
yansıtır. Bir doğal suş
(genetik olarak normal ve orijinal özelliklerini taşıyan bir virus) ile infekte edilen
bir canlıda oluşan antikorların etkinliği ile, bu virustan mutasyonla oluşturulan ve
değişik antijenik karaktere sahip mutant virusla infekte edilen canlıda oluşan
antikorların kros aktivite ve kros proteksiyon bakımlarından farklar
bulunmaktadır. Diğer bir ifade ile mutant suşa karşı oluşan antikorlar, orijinal
virusu belli bir oranda nötralize edebilir (veya bazen hiç nötralizasyon
görülmeyebilir). Ayrıca, böyle mutant suşlar da canlılarda, orijinal virus kadar bir
koruma sağlamayabilirler. Bu durum aynı zamanda, yerel suşlarla yapılan aşıların
(monovalan veya polivalan) kullanılmasının gerekli olacağını da ortaya koymaktadır.
Influenza ve retroviruslarında genetik varyasyonlara fazla rastlanıldığından,
bunlarda alt serotipler de ortaya sıkça çıkmaktadır. Retroviruslarında
replikasyonda 2 farklı enzim görev yapmaktadır. Bunlardan biri virionda kodlanan revers transkriptaz enzimi (RNA'ya bağımlı
DNA polimeraz enzimi) ile hücresel RNA polimeraz
(DNA'ya bağımlı RNA polimeraz) enzimleridir. Bunların da her ikisinin hata düzeltme
yetenekleri bulunmamaktadır. Bu durum da, retroviruslar arasında çok fazla mutant
suşların ortaya çıkmasına yol açmaktadır. Eğer mutasyon nukleik
asitlerdeki tripletlerin birinde, özellikle bir kodonun 3. bazında, ise sonuçta oluşan
değişiklikler proteinlerin yapısına pek fazla etkilemeyebilir, dolayısıyla da
önemli bozukluklar (proteinin aktivasyon noksanlığı veya inaktivasyonu) meydana gelmez
ve kısa süre içinde tamir edilebilir. Ancak, eğer üçüncü bazın değişmesi sonu
bir stop kodonu veya ardışık çok değişik kodonlar ortaya çıkıyorsa, o zaman
böyle mutasyonlar önemli olabilirler. Buna karşılık, kromozomal nitelik taşıyan
mutasyonlarda, (özellikle, delesyon tarzındakiler) çok önemli genleri taşıyan
segmentler çıkmışsa, virus inaktive olabilir ve letal mutasyon görülebilir. Virusların infeksiyonu
sonucunda hücrelerde ortaya çıkan plaklarda morfolojik değişiklikler
gözlemleniyorsa, bu farklı görünümdeki plaklardan ayrı ayrı elde edilen ve saf
olarak üretilen viruslardan hazırlanan aşılar deneme hayvanlarına verilerek oluşan
antikorların, doğal veya deneysel infeksiyonları önleme yönünden değerlendirilmesi
yapılarak, en uygun bulunanı aşı olarak kullanılabilir (Örn, Newcastle hastalığı
için hazırlanan Clone 30 aşısı gibi).
Ancak, böyle aşıların saha patojenik virusunu önlemedeki etkinliği, genellikle,
sınırlıdır. Bir gendeki mutasyonlar sonucu ortaya çıkan genotipik varyasyonlar, aynı yerde (değişikliğin olduğu yerde) meydana gelen geri mutasyonlarla veya aynı genin başka bir yerinde (intragenik) veya başka bir gende (ekstragenik) oluşan supresör mutasyonlarla giderilebilir. Mutasyonların en önemli
özelliği, baz sıralarında oluşan değişikliklerle genetik informasyonların (veya
tripletlerin değişerek başka amino asit kodonlarının sıraya girmesi ve bunun
sonucunda proteinin yapısında çok değişik amino asitlerin sırada bulunması) normal
düzeninin bozulmasına yol açmasıdır. 01.02. Başlıca Mutant Türleri Genlerin baz
sıralarında veya tripletlerde, çeşitli nedenlerle (spontan, fiziksel, kimyasal ve
diğer türden) meydana gelen genotipik değişmeler (mutasyonlar), virusun orijinal
(doğal) suşlarından farklı özellik taşıyan mutantların ortaya çıkmasına yol
açar. Bunlar arasında en önemli olan mutantlar aşağıda belirtilmiştir. 1) Özel koşullara bağlı letal mutantlar: Bazı
şartlara bağımlı olarak ortaya çıkan bu tür mutantlar, nonpermisif (uygun olmayan) koşullarda
infeksiyöz bir karakter taşımazlar. Ancak bazı özel uygun şartlarda (permisif koşullar) infeksiyöz projeniler
(nesiller) meydana getirmektedirler. Bu karaktere sahip mutantlar da başlıca iki
kategoride incelenmektedirler. a) Isıya duyarlı (temperatür sensitif, Ts) mutantlar:
Hayvan virusları arasında çok sık rastlanılan bu tür mutantlar, düşük ısıda
üremelerine karşın yüksek ısıda üreyememektedirler. Böyle mutantların
kodladıkları proteinler veya polipeptidler ancak düşük ısıda (30-31°C) aktivite
göstermekte, buna karşın daha yüksek ısılarda (37-38°C) etkinlikleri olmamakta ve
inaktif duruma gelmektedirler. Çünkü, yüksek ısı proteinlerin quarterner yapısını
bozmakta ve böylece aktivitesine olumsuz etkilemektedir. Viral genomda oluşan nokta
mutasyonlar, hücre içinde oluşturulan polipeptidlerde normal sırasında olmayan amino
asitlerin yer alması ve bunların da farklı karaktere (düşük ısıda aktivasyon
göstermesi gibi) sahip olmasına yol açmaktadır. b) Konağa bağımlı mutantlar: Böyle özellikteki
mutantlar ancak bir tür hücrede üreyebilir ve bunlarda plaklar oluşturabilirler. Buna
karşın bazıları da diğer bir türü tercih ederler. Poxviruslarında rastlanan p-mutantları ve adenoviruslarda saptanan kb-mutantları böyle özellik taşırlar. 2) Defektif mutantlar: Viral replikasyonları için
önemli olan bir veya bir kaç fonksiyonel geni bulunmayan viruslar, genellikle, bu
tanımla belirtilmektedirler. Böyle virusların duyarlı hücrelerde üreyebilmeleri
için, normal yardımcı (helper) viruslarının sağladığı
ve fakat, defektiflerin gen noksanlığı nedeniyle kendilerinin kodlayamadığı
proteinlere gereksinimleri vardır. Diğer bir ifade ile defektif viruslar, yalnız
olarak, permisif hücrelerde üreyemezler ve olgunlaşamazlar. Kendilerinde olmayan ve
fakat, normal viruslarca kodlanan gen ürünü proteinlere bağımlıdırlar. Böyle
durumlara delesyon mutantlarında fazlaca rastlanılmaktadır. Örn, defektif bir karakter
gösteren parvovirusların üreyebilmeleri için, hücrelerde normal (helper)
adenoviruslarının da bulunması gereklidir. Bazı durumlarda
virusların üremeleri, hücresel orijinli proteinlerin azlığından (yetersizliğinden),
aktivite noksanlığından veya yokluğundan da kaynaklanabilir. Laboratuvarlarda fazla
pasajı yapılan viruslarda defektif viral partikül sayısında artmalar
gözlemlenmektedir. Bunlar, ayrıca, homolog virusların da üremelerini
önleyebilmektedirler. Böyle mutantların, çoğu zaman infeksiyöz yetenekleri de
bulunmamaktadır. Ayrıca her hücre pasajında, infeksiyöz titrede azalmalar da
saptanır. Normal homolog virusların üremelerini inhibe eden böyle mutant viruslara "defektif interfere edici viruslar"
adı verilmektedir. Bu tür mutant virus partiküllerine RNA virusları arasında fazlaca
rastlanılmakta ve bunlar, aynı zamanda, persistent infeksiyonların ortaya çıkmasında
ve devamında da etkili olabileceği açıklanmaktadır. Eğer virusların üretilmeleri
için kullanılacak inokulumlar yüksek sulandırmalardan
hazırlanırsa (10-5 veya 10-6 gibi) defektif virus sayısında ve
hücrelerde birikmelerinde azalmalar meydana gelir. Defektif virusların
kapsidleri normal görünümde olmasına karşın genomlarında bir veya birkaç gen
noksandır. Halbuki, pseudoviruslarda ise,
kapsid normal olmasına karşın, virionda, viral genom yanı sıra konakcı hücre
DNA'sından da küçük bir segment bulunmaktadır. Bu nedenle de her ikisi de infeksiyöz
yeteneğe sahip değildirler. Ancak, pseudoviruslarda, genomun hepsi hücre orijinli
değildir. Defektif viruslara,
influenza virusları, adenovirusları, papova virusları ve diğer viruslar arasında
rastlanılmaktadır. 3) Sıcak mutantlar: Bu tür mutant viruslar, doğal
(orijinal) virusların ürediği ısı limitlerinin daha üstündeki derecelerde
(40-41°C) çoğalmaktadırlar. Böyle mutantlar, vücut ısıları (ateşi) yükselmiş
hastalarda kolayca üreyebilmekte ve infeksiyonlara yol açabilmektedirler. 4) Plak mutantları: Aynı virus türlerinde,
spontan veya çeşitli faktörlerle oluşturulan mutasyonlar sonunda, plak formasyonlarında da bazı morfolojik
değişiklikler de meydana gelmektedir. Böyle mutantlar genellikle delesyonlar sonucunda
ortaya çıkmaktadırlar. Aynı tür içinde heterolog plakların görülmesi antijenik ve
patojenik karakterlerindeki değişmelerden kaynaklanmaktadır. Plak pürifikasyonları
ile saf virus partikülleri elde edilebilir. 5) İlaçlara dirençli mutantlar: Bazı ilaçlara
direnç gösteren mutantlara viruslar arasında tesadüf edilmektedir. Örn, poliovirus
mutantının guanidine ve bazı herpes
virus mutantları da fosfoasetik asite
direnç gösterdikleri belirtilmiştir. Bunlara karşın, polioviruslarında ve vaccinia
viruslarında guanidine bağımlı
suşların varlığı açıklanmıştır. Viruslar arasında
bazı fiziksel ve kimyasal maddelere (formol, vs) dirençli nesillerin meydana geldiği de
bildirilmiştir. 02. Rekombinasyonlar 02.01. Genel Bilgiler İn vivo veya in vitro
(hücre kültürlerinde) üreyen virusların arasında veya viruslarla içinde üredikleri
hücre DNA'sı arasında oluşan genetik
interaksiyonlar sonunda karşılıklı
genetik materyal değişimi (intramoleküler genetik değişim), genellikle, rekombinasyon olarak tanımlanmaktadır. Bu olgu
sonunda da yeni genetik karakterlerde hibrid
virus suşları oluşmaktadır. Bu suşlar, doğal viruslardan bazı yönleri ile
farklı oldukları gibi kendisini oluşturan her iki virusa ait genetik materyallere de
sahiptirler.
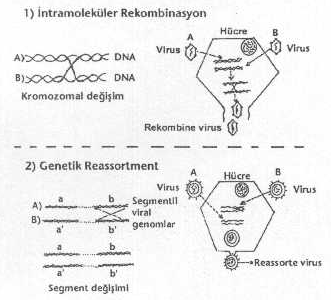
1) İntramoleküler rekombinasyonlar: Bir hücre,
genetik olarak birbirine yakın iki virusla koinfekte
edildiğinde (örn, HSV-1 ve HSV-2 virusları gibi) viral nukleik asitlerinin sentezleri
sırasında karşılıklı gen alış-verişi meydana gelmekte ve oluşan yeni hibrid suşlar, hem tip-1 ve hem de tip-2
HSV'larına ait genetik materyallere sahip olmaktadırlar (intertipik rekombinant viruslar). Diğer DNA
viruslarında da (SV-40 ile adenoviruslar) benzer rekombinasyonlar ortaya
çıkmaktadırlar. 2) Genetik reassorment: Bu olgu daha ziyade
segmentli viruslar arasında görülmektedir. Aynı hücre, genetik olarak birbirine çok
yakın iki virus tarafından koinfekte edildiğinde bu iki virusa ait bazı segmentler
arasında karşılıklı değişmeler (segment
değişimi) olmaktadır. Bu fenomen sonunda da hibrid virus partikülleri meydana
gelir. Bu olayda, her iki virusa ait segmentler karşılıklı bir araya (yan yana)
geldiklerinde segment değişimi görülür. Bu değişim sonunda oluşan
reassortmentlerden aşı suşu olarak kullanılabilir. Ancak, hepsi bu amaç için uygun
değildir ve çok iyi bir seleksiyon yapmak gereklidir. Reassorment tarzındaki
rekombinasyonlara influenzaviruslarında sıkça rastlanmakta ve virusun, doğada, 13 tür
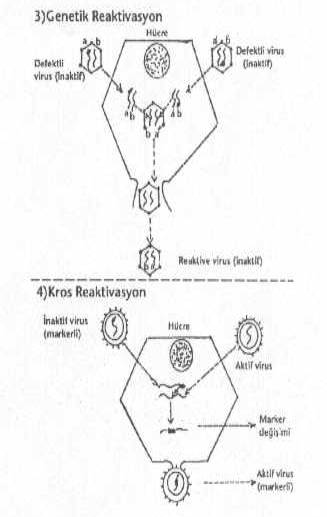
hemaglutinin H1-H13 ve 9 türde neuraminidaz varyantı bulunmaktadır. 3) Genetik reaktivasyon: Bir hücre, farklı
genlerinde defekt bulunan ve birbirine genetik çok yakın (akraba) iki inaktif virus tarafından infekte
edildiğinde, hücre içinde infeksiyöz virus partiküllerinin oluşmasına multiplisite reaktivasyonu adı verilir. Böyle
olgular UV-ışınları ile inaktive edilen viruslarda daha iyi gözlenmektedirler.
Kimyasal maddeler de benzer tarzda etkili olabilmektedir. 4) Eğer bir hücre
biri aktif ve diğeri de inaktif birbirine genetik yakın olan iki virus tarafından
infekte edilirse, aralarında, hücre içinde gen alışverişi olabilmektedir (kros reaktivasyon, Marker rescue). Bu olguda,
inaktif virusun genom porsiyonu, aktif virusunun genomuna rekombine olmaktadır. Böylece
ortaya çıkan yeni projeniler, genomlarında inaktif virusa ait özel markerleri
taşırlar.
02.02. Viral Gen Ürünleri Arasında İnteraksiyonlar Viruslar arasında
genetik olmayan bu tarz interaksiyonlar bir kaç şekilde ortaya çıkmaktadır .
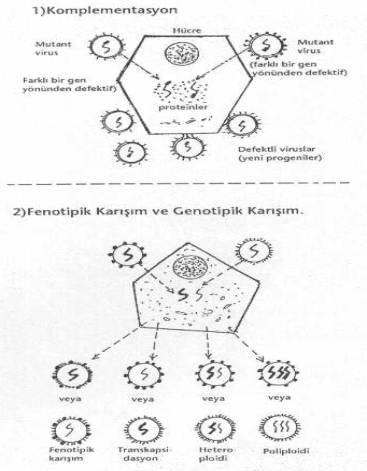
1) Komplementasyon: Her biri yalnız olarak
hücrelerde üremeyen iki mutant virus aynı hücreyi infekte ettiklerinde birinin
replikasyonu için gerekli olan proteinin diğeri kodlayarak her ikisi birlikte aynı
hücrede üreyebilirler. Komplementasyon, birbiri ile ilişkili olmayan viruslar, defektif
viruslar veya zarflı-zarfsız viruslar arasında görülebilir. Bu olguda, belli bir gen
yönünden defektif olmayan bir virusun sağladığı gen ürünü, bu ürünün sentezini
kodlamayan defektif virus tarafından kullanılarak kolayca üreyebilmektedir. Bu iki
mutant virus arasında gen değişimi olmadığı için, yeni projenilerde de defektif
genler bulunmaktadır. Eğer, iki mutant virus, aynı ürünün geni için defektif iseler
o zaman her ikisi de üreyemez . 2) Fenotipik karışım: Bir hücrenin miks
infeksiyonu sonu, bir virusa ait genetik materyalin tümü, diğer virusa ait kapsid veya
zarfın içine girebilir (transkapsidasyon).
Bazen de, bir virusun kapsid veya zarf materyalleri, diğer virusa ait kapsid veya zarf
proteinleri arasında bulunabilirler (fenotipik
karışım). Bu son durumda, viral genom etrafında her iki virusa ait proteinden
meydana gelen kapsid yer alır. 3) Poliploidi ve heteroploidi (genotipik karışım): Virusların hücrelerde replikasyonları sırasında bazen aynı zarf veya kapsid içinde birden fazla aynı virusun genetik materyali (poliploidi) veya bir kapsid veya zarf içinde değişik viruslara ait genetik materyaller bulunabilirler (heteroploidi). Bu tarzdaki olgulara, daha ziyade, hücrelerden tomurcuklanma ile çıkan viruslarda (paramyxoviruslar ve diğerleri) görülmektedir. |
||||||