|
Translasyon (Protein Biyosentezi) Prof. Dr. Mustafa Arda
Ankara Üniversitesi
Veteriner Fakültesi 01. Genel Giriş 01.Genel Giriş Proteinler,
hücrelerin informasyonal makromolekülleri olup kromozomda bulunan nukleotid (baz)
sıraları tarafından spesifiye edilirler. Her protein bir veya birden fazla polipeptid
zincirinden oluşmuştur. Her bir peptid zinciri de, aminoasitlerden meydana gelmiş
lineer polimerler halindedirler. Proteinlerin amino asit sıraları, mRNA'daki baz
sıraları tarafından tayin edilir. Genellikle, bir genin bir proteini kodladığı kabul
edilir. Mesenger
RNA'daki kodlar, ribosomlar üzerinde deşifre edilerek, amino asit sıraları (polipeptid
ve proteinler) haline dönüştürülürler Protein
sentezi bilinen en kompleks biyokimyasal olaylardan biri olup 100 den fazla farklı
protein ve 30 türden fazla tRNA'nın fonksiyonu ile gerçekleştirilir. Protein
sentezindeki sellüler mekanizmaları kolay izleyebilmek ve anlamak için, bunları
aşamalar halinde incelemek daha yararlı olacaktır. Protein
sentezi, başlıca 4 basamak içinde toplanabilir; 1)
Aktivasyon 2)
Başlama 3)
Zincir uzaması 4)
Bitiş (terminasyon) 02. Aktivasyon Hücre
içinde, sitosolda bulunan amino asitler, mRNA üzerindeki spesifik kodonlarına
taşınmadan önce, kendine ait spesifik tRNA ile interaksiyona girmesi ve bağlanması
gereklidir. Her amino asitin kendine uygun bir
veya birden fazla tRNA'sı bulunmaktadır. Amino asitleri mRNA'ya taşımada
tRNA'lar adaptör molekül görevi yaparlar. Amino asitlerin, kendilerine uyan tRNA'lara
bağlanmasında amino acyl synthetase enzimi önemli
fonksiyona sahiptir ve amino asitleri aktive ederek, tRNA'ya bağlanmasını sağlar.
Aktive olan amino asitler, kendilerindeki karboksil grubu ile tRNA'nın serbest 3'-CCA
(adenilik asit) terminusuna (akseptör kol) ester
bağlarıyla birleşir. Reaksiyon iki aşamada tamamlanır. Gerekli enerji ATP'den
sağlanır ve reaksiyonu sentetase enzimi katalize eder. Sonuçta,
Mg++ Amino
asit + ATP + tRNA ¾® amino acyl tRNA + AMP + PP1 meydana gelir. Her
sentetase enzimi, hem spesifik amino asitin ve hem de ilgili tRNA'nın üzerindeki özel
bölgeleri tanıma yeteneğine sahiptir. Hücrelerde, genetik kodlar tarafından spesifiye
edilen her bir amino asit için bir tane sentetase enziminin bulunduğu bildirilmektedir.
Ancak, enzim, birden fazla ilgili tRNA'yı tanıma yeteneğindedir. Hatta, enzim,
tRNA'ların antikodonları değişse veya çıkarılsa bile yine de (uygun) tRNA'yı
tanıyabilir. Fakat, bazı sentetazlar ise hiçbir değişikliğe maruz kalmamış
antikodonları tanıyabilmektedir. Birçok
sentetase enziminde hataları düzeltmek için bazı spesifik bölgelerin bulunduğu
belirtilmiştir. tRNA'lara uygun olmayan amino asitlerin bağlanması durumunda, enzim,
bunları hidrolize ederek çıkarır ve doğru amino asitle yer değişmesini sağlar (geriye dönük düzeltme
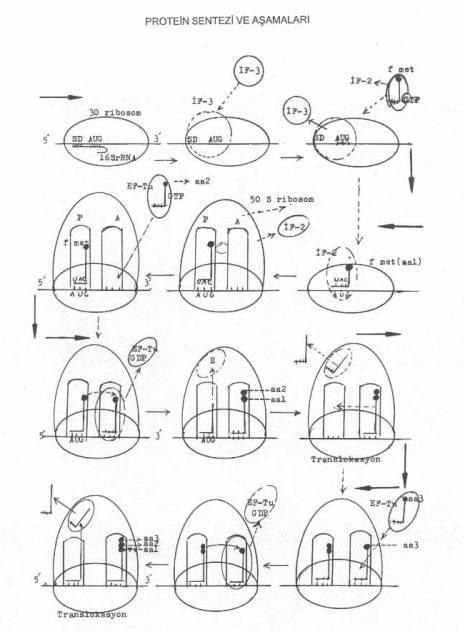
okuması). 03. Başlama Protein
sentezini başlatmada özel tek bir tRNA görev yapar ki o da formilize olmuş metionini taşıyan fmet tRNA'dır. Bu transfer RNA, aynı zamanda,
kendi antikodon bölgesindeki bazlar (UAC)
yardımıyla, mRNA üzerinde sırada bulunan ve metioninin kodonu olan AUG'yi tanıyarak kovalent olarak bağlanır.
Bakterilerde, bazen AUG yerine, nadiren GUG kodonu da kullanılabilir.
Hücrelerde,
genellikle, iki tür tRNA, metionini spesifiye eder. Bunlardan biri, internal pozisyonda
bulunan metionini tanır ve onunla bağlanır (met
tRNA). Diğeri ise, formilize olmuş metioninle bağ
kurar (fmet tRNA). Ancak, bunlardan sadece fmet tRNA başlatmada rol alır ve önem taşır. Aynı zamanda, başlatma faktörleri (IF), fmet tRNA'yı, ve uzatma faktörleri de (EF), met tRNA'ları tanırlar. Başlatmada
diğer önemli aşama ise, formilize olmuş metioninin tRNA'sının antikodonu (UAC) ile,
30 S'lik ribosomal alt ünite üzerinde sıraya girmiş olan mRNA'daki kendine ait AUG
kodonunun bağlanmasıdır. 30 S
ribosomal serbest alt ünite önce başlatma faktörü (IF-3) ile bağlanır. Bu faktör,
30 S'lik alt ünitenin, mRNA'nın 5' -ucuna ve AUG kodonuna bağlanmasını güven altına
alır. IF-3 olmadan 30 S alt ünite mRNA ile
bağlanamaz. Böylece, 30 S'lik alt ünite mRNA'nın 5' -ucundaki AUG kodonu ile
bağlanmış ve protein sentezi için güvenli ilk adım atılmış olur. Bu aşamadan
sonra, başlatma faktörlerinden olan IF-2 fmet
tRNA ile bağlanarak bunu, ribosoma getirmede görev alır. Bununla beraber,
reaksiyonda gerekli olan enerjiyi sağlayacak olan GTP'de ribosoma getirilir. fmet tRNA
gelmeden ve 50 S'de birleşmeden önce, görevi sona eren IF3 ayrılır ve hidrolize olan
GTP'de GDP'ye dönüştürülür. mRNA,
30 S'lik küçük alt ünite ile birleştikten sonra IF-3 hemen ayrılır. 30 S alt
ünitede P-bölgesinde sırada bulunan ve IF-2 ile bağlanmış olan fmet tRNA'nın
antikodon bölgesindeki bazlar (UAC), mRNA'da sırada bulunan kendine ait olan kodon (AUG)
ile birleşir. Bu iki bölge birbirine komplementerdir. mRNA'da
AUG kodonunda 5' -ucuna doğru ve bundan yaklaşık 10 baz kadar uzakta pürinden zengin
olan Shine Dalgarno (SD) sekansları da
mRNA'nın ribosomlara sıkıca bağlanmasında rolü vardır. Shine Dalgarno bölgesindeki
bazlar ile 30 S'lik alt ünitede bulunan 16 S rRNA'daki nukleotidler birbirlerine
komplementerdir. Böylece kolay bir bağlanma yaparlar. Bu sekanslar, mRNA'nın ve
dolayısıyla da mesajların daha güvenli ve sağlam bağlanmasını sağlarlar. Prokaryotiklerde
DNA üzerinde başlatma kodonundan 3' -ucuna doğru, promotor bölgesi de bulunmasına
karşın, mRNA'ya sadece SD ve (DNA da
bulunan TAC tripletinin karşılığı olan) AUG
bazları aktarılır, promotor aktarılmaz. 30
S'lik alt üniteye fmet tRNA bağlandıktan hemen sonra 50 S ribosomal büyük alt ünite
de katılır. Böylece başlama kompleksi oluşur
(70 S). Hemen IF-2 (ve IF-1) kompleksten çıkarılır. GTP'de hidrolize olur ve
GDP + PP1'e ayrışır. 50
S'lik ribosomal alt ünitede bulunan bölgelerden P-bölgesinde (peptidil bölgesi) fmet tRNA lokalize olmuştur
ve A-bölgesi (akseptör bölge) ise boştur. Ancak,
burada polipeptid zincirine girecek olan ikinci amino asitin kodonu bulunmaktadır.
İkinci amino asitin (aa2) sıraya girebilmesi için, sentetase enziminin yardımı ile
kendine ait tRNA ile birleşmesi ve bu defa zincir
uzatma faktörüne (EF-Tu) bağlanarak A-bölgesinde hazır bulunan mRNA
üzerindeki kendine ait kodona, antikodonuyla birleşmesi gerekir. EF-Tu+amino
asil tRNA+GTP, üçlü kompleksi ribosom üzerinde A-bölgesinde sıradaki kodona,
tRNA'nın antikodon bölgesindeki nukleotidler ile bağlanır. Bağlanma bitince EF-Tu
serbest kalır. GTP'de GDP'ye ve PP1'e ayrışır. Böylece,
P-bölgesinde fmet tRNA (1. amino asit, aa1) ve A-bölgesini de 2. amino asiti (aa2)
taşıyan tRNA (amino asil tRNA) işgal etmiş olur. Bundan sonra, P-bölgesindeki fmet
ile, A-bölgesinde tRNA'ya bağlı bulunan 2. amino asit arasında peptid bağı kurularak
P bölgesindeki aa1 (fmet) A-bölgesine transfer edilir ve aa1 ile aa2 birleşir. Bu
suretle, A-bölgesinde tRNA'nın 3' -CCA ucunda iki amino asit bağlı bulunmuş olur.
İki amino asit arasında peptid bağının
kurulmasını peptidil transferase enzimi katalize eder. Bu suretle,
polipeptid zincirine iki amino asit katılmış olur. P-bölgesinde serbest kalan f metionine ait tRNA buradan, geçici bir süre
için E-bölgesine transfer edilir ve sonra
da buradan ayrılır. Her
bakteride yaklaşık EF-Tu'den 70000 molekülün bulunduğu belirtilmiştir. Diğer bir
zincir uzatma faktörü olan EF-Ts, EF-Tu+GDP inaktif molekülünü aktive ederek tekrar
EF-Tu+GTP'ye çevirir. EF-Tu+EF-Ts'lere T-faktörleri adı verilir. 04. Zincir Uzaması Başlatma
kompleksi oluştuktan sonra ikinci amino asitin kodonla birleşmesi, zincir uzamasının
ilk adımını da oluşturur. Bu periyot başlıca 3 kısımdan meydana gelir. a)
Kodon tanıma: Bu
aşamada, yukarda belirtildiği gibi mRNA üzerinde sıraya giren ve üç bazdan oluşan
kodonla bu kodonun temsil ettiği amino asitle birleşen tRNA'nın antikodon bölgesi
arasında, bunların komplementer olması nedeniyle, karşılıklı bağlar kurulur. Örn,
eğer kodon üzerinde Valinin tripletinden biri olan GUU varsa, buna tekabül eden ve valini taşıyan
amino acyl tRNA'nın antikodonunda CAA
bazlarının bulunması gerekecektir. Böylece,
tRNA'ya bağlanmış olan amino asitte, EF-Tu+GTP ile birlikte, 30 S ribosom alt
ünitedeki kodonla birleşir. Birleşme olunca EF-Tu serbest kalır, GTP'de, GDP ve PP
veya 1'e ayrışır (EF-Tu-GDP inaktif bir moleküldür). b)
Translokasyon: 70
S'lik ribosomda P- ve E-bölgeleri boşaldıktan sonra, A-bölgesinin de boşalması ve
buraya 3. amino asiti bağlayan tRNA'nın gelmesi gerekir. Bunun için, A-bölgesinin
serbest kalması lazımdır. İşte bu önemli olay, 70 S'lik ribosomun mRNA üzerinde bir
kodon boyu kayması (5' ® 3'
yönde) ile gerçekleşir. 70 S'lik ribosom, m RNA üzerinde sadece bir kodon boyu
kaydığında. A bölgesi, m RNA üzerindeki bitişik, yeni bir amino asitin kodonuna
gelmiş olur (veya böylece, A bölgesine yeni bir kodon gelmiş olur). Böyle bir hareket
olurken, P bölgesindeki AUG kodonuna bağlı olan ve amino asit taşımayan t RNA
dışarı itilerek serbest kalır ve AUG kodonundan ayrılır. Serbest kalan tRNA önce 50
S alt ünite üzerindeki E bölgesine gelir ve buradan da diğer bir amino asitle
bağlanmak için tekrar sitosola döner. Bu kodona ait amino asitleri, amino acyl sentetase aktive ederek kendine ait
tRNA ile bağlar. Bundan sonra, EF-Tu ve GTP ile bağlanan amino acyl tRNA kompleksi,
kodonla birleşir. Translokasyonda
ribosom, mRNA üzerinde her defasında bir kodon boyu ilerler. Translokasyon için
başlıca 2 görüş bulunmaktadır. Biri, önce 50 S'lik alt ünite 5' ® 3'
yönünde 30 S'in üzerinde bir kodon boyu kayar ve sonra bunu 30 S'lik alt ünite takip
ederek 70 S'lik ünite tamamlanır. 50 S'lik alt ünitenin kayma sırasında, 30 S
üzerindeki P-bölgesindeki peptidil tRNA'lar ile A-bölgesindeki amino acyl tRNA'lar
yerinde kalırlar. Diğer görüş ise, 70 S'lik ünite tümden mRNA üzerinde bir kodon
boyu ilerler. Translokasyonda
başlıca 2 faktör önemlidir. Bunlardan biri, EF-G ve diğeri de GTP'dir. EF-G her
hücrede bir kopya halinde bulunur (20.000 molekül). EF-G, ribosoma bağlanarak
translokasyonu güven altına alır. GTP hidrolize olunca, EF-G'de serbest kalır. Ribosomlar,
aynı anda, hem EF-Tu ve hem de EF-G ile ilişkili kuramazlar. Bu faktörler ribosomlara
sıra ile bağlanırlar. Birinin fonksiyonu bitip kompleksten ayrılırken diğeri,
reaksiyona katılır. Önce, EF-Tu+GTP+amino acyl tRNA ternar (üçlü) kompleks oluşturur. Bundan
EF-Tu+GDP ayrıldıktan sonra EF-G+GTP bağlanır ve sonra da EF-G+GDP halinde serbest
kalır. c)
Transpeptidasyon:
Transpeptidasyon, peptidil bölgesindeki tRNA'ya bağlı amino asitler ile
A-bölgesindeki, yeni gelen, tRNA'daki amino asitin karboksil ve amino terminal uçları
arasında peptid bağı kurulması olayıdır. Bu reaksiyonu peptidil transferase enzimi katalize eder. Bu
reaksiyon sonunda, P-bölgesindeki amino asitler, A-bölgesindeki yeni gelen amino asitle
birleşirler. Böylece
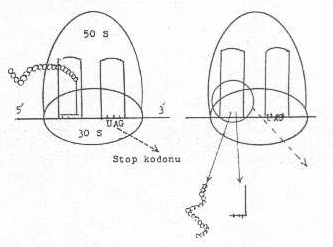
zincir uzaması devam eder. Her translokasyonda sıraya bir amino asit katılmış olur. 05. Bitiş (terminasyon) Protein
sentezini sonlandırmada mRNA üzerinde 3 kodon etkili olur ve bunlara terminasyon kodonları (stop kodon) adı verilir (UAG: amber; UAA: ochre ve UGA:opal). Bazen terminasyon tripletlerini ifade
etmede nonsense kodonlar terimi de
kullanılmaktadır. Bakteriler,
en fazla UAA tripletini terminasyon kodonu olarak kullanırlar. Diğer terminasyon
kodonlarından daha az yararlanılır. Eğer,
mRNA üzerinde bulunan bu kodonlardan biri A-bölgesine gelirse, polipeptid zincirine
herhangi bir amino asit ilave edilemez. Çünkü, bu kodonların karşılığı olan
spesifik bir amino asit ve tRNA yoktur. Böylece sentez durur. Fakat, polipeptid henüz
mRNA'dan ayrılmış değildir. E. coli 'de
zincirinin ayrılmasında 2 proteinin rolü bulunmaktadır. Bunlardan biri, serbest bırakma faktörü-1 (RF-1) ve diğeri de
RF-2'dir. Bunlar farklı terminasyon kodonu
için spesifiktirler. RF-1, UAA ve UAG'yı tanır, RF-2 ise UGA ve UAA'yı tanır. Bu
faktörlerden birisi terminasyon kodonuna geldiğinde polipeptid zinciri tRNA'dan serbest
kalır. Polipeptidin tRNA'dan ayrılması, moleküle suyun ilavesi ile (hidrolizasyon) gerçekleştirilir. Her bir
hücrede bu faktörlerden yaklaşık, 600 kadar bulunur. Diğer bir serbest bırakma faktörü (RF-3) olarak tanımlanan proteinin, diğer iki
faktörünün (RF-1 ve -2) aktivitesini kuvvetlendirdiği bildirilmektedir.
Protein
sentezi sona erdikten ve protein serbest kaldıktan sonra iki alt ünite de (30 S ve 50 S)
birbirinden ayrılarak sitosola dönerler. 06. Önemli Notlar -tRNA'lardaki
antikodon (lar) ile mRNA'daki kodon(lar) birbirlerinin komplementeridirler. -Prokaryotiklerde
ekspresyon (transkripsiyon ve translasyon) sitoplazmada, buna karşın ökaryotiklerde
transkripsiyon nukleusta ve translasyon ise sitoplazmada meydana gelir. -Proteinler,
ribosomlar üzerinde N-terminustan C- terminusa doğru asamble edilirler. -Ribosomlar,
mRNA üzerinde 5' ¾®3'
yönünde hareket ederler. Her bir hareket bir kolon boyudur ve her defasında bir yeni
amino asitin kodonu sıraya girer. -Her
ribosom, mRNA boyunca bağımsız olarak bir polipeptidin sentezini gerçekleştirebilir. -mRNA,
aynı anda, birçok(onlarca) ribosom tarafından transle edilebilir. -mRNA'ya
bağlanan ribosomların bir kısmı sentezin sonunda iken diğeri başlangıcında
olabilir. -Antikodon
ve kodon ilişkisinde ilk iki nukleotid (baz) önemlidir. Üçüncü bazın değişikliği
çok fazla etkili olmayabilir. Yani, 3. baz, kodlanan amino asit türü değişmeden,
değişik türde olabilir. -Kodonlar
(tripletler) bir biri arkasına dizilmiştir. Bakteriler de aralarında başka veya kodlamayan sekanslar (intron) bulunmaz.
Bakterilerde DNA veya RNA ekzonlardan (kodlayan sekanslar) oluşur. Ökaryotiklerde genlerin içinde ve aralarında
kodlamayan sekanslar (intron) vardır. -Amino
asitlerin, kendilerine ait tRNA'larına bağlanmalarını, amino acyl tRNA synthetase enzimi katalize eder.
En azından 20 sentetase enzimi vardır ve her biri ayrı amino asiti tanır. -Prokaryotiklerde
mRNA, genellikle, polisistroniktir (birden fazla polipeptid
zincirini kodlar), ökaryotiklerde monosistroniktir. -Bir
amino asit bir veya birden fazla sinonim kodonla temsil edilebilir. -Genetik
kodlar üniversal olmasına karşın, azda olsa bazı değişimlere rastlanmaktadır.
Örn, mycoplasmalarda UGA kodonu tryptophanı kodlamasına karşın, diğer
prokaryotiklerde bu triplet stop sinyalini
verir. -Protein
sentezi bakterilerde çok çabuk oluşan ve aynı zamanda ısıya bağımlı bir
prosestir. Bakteriler 37°C 'de, saniyede yaklaşık 15 amino asiti polipeptid zincirine
katar (300 amino asitlik bir protein için yaklaşık 20 saniye yeterli olabilmektedir).
Ökaryotiklerde protein sentezi çok daha yavaş ilerler (saniyede 1-2 amino asit
ilavesi). -IF-3
taşımayan 30 S alt ünite, mRNA ile bağlanamaz ve başlatma kompleksi oluşamaz. 70
S'lik ribosomda IF-3 bulunmaz. -IF-3,
30 S'den ayrılınca, bu defa başka ve serbest 30 S alt ünite ile bağlanır. -Her
bakteride yaklaşık 70000 molekül EF-Tu bulunur. İki gen tarafından spesifiye edilen
EF-Tu, bakteri total proteininin %5'ini oluşturur. Buna karşın Ts ise yaklaşık 10000
molekül kadardır. -IF-3,
mRNA'nın 50 S ile bağlanmasını önler. 07. Translasyon İnhibitörleri Mesenger
RNA molekülünün translasyonunda bir veya birkaç basamağı inhibe ederek proteinin
biyosentezine mani olan bir çok substans bulunmaktadır. Bakterilerdeki
protein sentezi üzerine etkili olmasına karşın ökaryotikler de tesirsiz olan bazı
antibiyotikler ve aktiviteleri aşağıda gösterilmiştir.
[1] Kaynak: Temel Mikrobiyoloji |
||||