|
Nukleik Asitlerin Biyolojik Fonksiyonları Prof. Dr. Mustafa Arda
Ankara Üniversitesi
Veteriner Fakültesi 01. Genel Giriş
Nukleik
asitlerin hücre içindeki en önemli görevlerinden birisi de, kuşkusuz, DNA'nın replikasyonu ve bu sayede genetik bilgilerin
nesillere aktarılması ile genlerin ekspresyonu (transkripsiyon ve translasyon)'dur. 02. DNA
Replikasyonu Hücreler,
tek kromozomlu (prokaryotik) veya çok kromozomlu (ökaryotik) olsunlar, DNA'lar genetik
bilgileri taşıdıklarından, her hücre bölünmesinde, bunların da çok az hata ile
replike olması ve yeni sentezlenen DNA'nın kardeş hücrelere eşit olarak aktarılması
gereklidir. Replikasyon tamamlanıncaya kadar da hücrelerin bölünmemesi lazımdır.
Aksi halde, bazı hücreler genetik materyallerden yoksun kalır ve çekirdeksiz hücreler oluşur (anukleer hücreler). Diğer bir önemli nokta da,
kromozomun (DNA), dış ve içten kaynaklanan zararlı etkilerden korunması ve DNA'da
meydana gelecek bozuklukların da hemen tamir edilerek nesillere aktarılmasının
önlenmesidir. Böyle olumsuz durumları gidermek için hücreler yeterli düzeltme ve
enzimatik mekanizmalara da sahiptirler. Genetik
materyallerin her ne kadar iyi korunmuş olmasına karşın, DNA'nın çok sayıda
replikasyonu ve çok değişik nedenler bazı mutasyonların meydana gelmesine yol açmakta ve buna
bağlı olarak da değişik karakterde yeni nesiller (mutant) ortaya çıkmaktadır. DNA'nın
hücre içinde kısa bir sürede ve minimal bir hata ile nasıl replike olduğuna dair
bazı görüşler ileri sürülmüştür. Bunlardan 1) Konservatif (parental iplikçikler birbirlerinden
ayrılmadan kopyalarının çıkması) ve Dispersif (parental DNA'da kopmalar meydana
gelerek aralarına yeni sentezlenen DNA segmentlerinin girmesi) olan teoriler terk
edilerek yerini, Watson-Crick tarafından ileri sürülen ve
araştırıcılar tarafından da (Meselson
ve Stahl deneyi, E. coli 'de) ispatlanan Semi konservatif modele bırakmıştır. Buna
göre, sarmal ve dubleks DNA açılmakta ve her iplikçiğin (parental iplikçik)
karşısında enzimler tarafından bir yenisi sentezlenmekte ve parental iplikçikler yeni
sentezlenen iplikçikler için bir kalıp görevi yapmaktadırlar. Bu modelde, her bir
çift iplikçikli DNA'da bir parental ve bir de yeni sentezlenen iplikçik bulunmaktadır. Prokaryotiklerde
(bakterilerde) ispatlanan bu semi konservatif replikasyon modelinin, ökaryotikler, virus
ve fajlar için de geçerli olduğu belirlenmiştir. Bu durum, semi konservatif
replikasyon tarzının genel (üniversal)
bir karakter taşıdığını da ortaya koymaktadır. 03. Prokaryotiklerde Replikasyon E.
coli
'ler
üzerinde çok fazla deneme yapıldığı için, bu mikroorganizma, replikasyon için de
bir model oluşturmuştur. E.
coli
genomu,
sirküler, sarmal ve dubleks bir DNA karakteri taşıyan ve yaklaşık 4.5x106 baz çiftine (bp, base pair) sahip tek bir
makro moleküldür (1.1-1.4 mm uzunlukta). Tüm genomun replikasyonu için 20-50 gen'in
fonksiyonel olduğu belirtilmektedir. E.
coli uygun koşullar altında 15-20 dk. bir generasyon meydana getirmektedir.
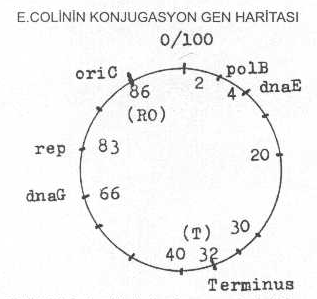
Genom replikasyonu E. coli 'de tek
bir orijinden (oriC) başlayarak bidireksiyonal (iki yöne doğru) olarak devam eder.
E. coli
'de
replikasyon orijini, konjugasyonla belirlenen dakika durumuna göre, 86. dk'dan iki yöne
doğru devam ederek 32. dakikada son bulur (terminus).
Bu süre, optimal koşullarda, bakterinin 15-20 dakikada bir bölünmesi süresinden daha
kısadır.
E.
coli
'de,
kromozomun replikasyon orijini 86. dk. bölgesi olmasına karşın, diğer bakterilerde
hem orijin ve hem de terminus daha değişik yerlerde olabilmektedir. Bu noktalar, bakteri
türlerine özeldir. E. coli 'de
bile uygun olmayan koşullarda bölünme süresi, optimal limitleri geçebilir. Replikasyonun,
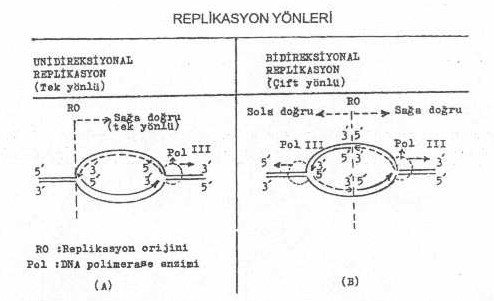
kromozomu küçük olan bazı mikroorganizmalarda, tek yönlü (unidireksiyonal) olduğu da
belirtilmiştir. Bu durumda, replikasyon, orijinden sadece bir yöne doğru devam eder ve
tekrar başlangıç yerinde son bulur. Tek
veya çift yönde replikasyonun olma durumu, orijin tarafından tayin edilir. Radyoaktif
işaretleme yöntemleriyle, bu özellik saptanabilir. E. coli 'de, replikasyon orijininde yaklaşık 245 bp
bulunmakta ve burada A-T bazlarının fazla olması açılmayı kolaylaştırmaktadır. E.
coli
'de
replikasyon fenomeninin başlangıcında genom sitoplasmik membranda bulunan özel bir
bölgeye (mesosom) bağlanır. Burası,
bakterinin yaklaşık orta bölgesine isabet eder. Diğer
önemli nokta, kromozomun, sitoplasmik membranda bağlantı yerine kuvvetlice
tutunmasıdır. Bunlara yardımcı olan bazı özel proteinler de belirlenmiştir. Genom,
özel bölgeye bağlandıktan sonra buraya replikasyonda görevi olan bazı spesifik
enzimler, proteinler ve dört tür deoksiribonukleotid trifosfat (dNTP) molekülleri
birikir. Bunlar arasında, DNA polimerase,
DNA ligase, helikase, primase, SSB, primosom,
DNA'yı
açan proteinler ve topoisomerase en önemlileri arasındadır.
Ayrıca, dNTP'ler ve Mg++ iyonlarına gereksinim vardır. Replikasyonu
bazı aşamalar halinde incelemek, olayı anlamak bakımından yararlı olacaktır. 1)
Replikasyon noktasının belirlenmesi:
E. coli 'de
kromozom üzerinde 86. dk.'ya isabet eden bölge replikasyon orijinini (oriC) oluşturur ve burası yaklaşık 245
bp'den meydana gelmiştir. DNA, sitoplasmik membrandaki bu özel bölgeyi çeşitli
proteinlerin yardımıyla da tanıyarak sıkıca bağlanır. 2)
DNA'nın açılması:
Bağlanma işlemi gerçekleştikten sonra, DNA'daki replikasyon orijini bölgesinde
küçük bir açılma meydana gelir. Bu açılmayı, özel açıcı proteinler (enzimler),
DNA iplikçikleri arasını açmak suretiyle genişletirler. Böylece sağa ve sola doğru
giderek açılan bir replikasyon çatalı
oluşur.
Başlangıç
bölgesinde DNA'da ilk açılmanın oluşu tam olarak aydınlatılmış değildir. Bir
çok bilgilere sahip olunmasına karşın yine de çözüm bekleyen bazı noktalar
bulunmaktadır. E. coli 'de 245
bp'den oluşan orijin bölgesinde kolayca açılabilen ve A-T bazlarından zengin 3 tane
13 bp ve 4 tane de 9 bp'lik tekrarlanan sekanslar vardır. Önce, dnaA proteinleri oriC bölgesinde toplanır ve
reaksiyon bu iki bölgede (9 bp ve 13 bp) başlatılır. İlk açıklık 13 bp.'lik
bölgede 3 yerden başlar ve sonra, dnaB
ve dnaC'nin de komplekse katılmasıyla
açıklıklar genişletilir ve bidireksiyonal
replikasyon çatalı meydana gelir. 3)
DNA sentezi (polimerizasyon): Parental
iplikçikte açılmanın meydana gelmesiyle oluşan replikasyon çatalında, sentez hemen
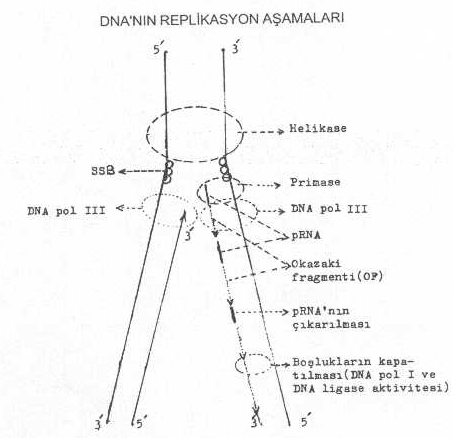
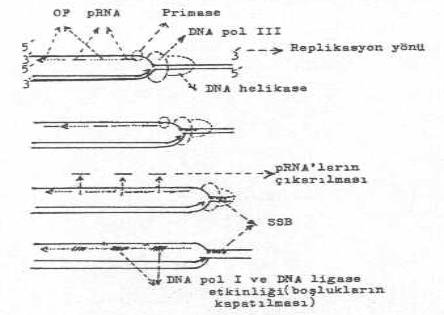
sağa ve sola doğru ilerlemeye başlar. Sentezi gerçekleştiren DNA polimerase-III
enziminin önünde iki önemli enzim daha fonksiyoneldir ve bunlar da sentez yönünde
ilerler. Bunlardan biri, eğer, DNA'da süper
heliks (super sarmal) varsa bunu açarak genomu istirahat haline getiren topoisomerase (DNA girase) enzimi ve diğeri de istirahat
halindeki DNA'da karşılıklı hidrojen bağlarını açarak, devamlı replikasyon
çatalı oluşturan helikase (dnaG)
enzimidir. Açılan iplikçiklerin tekrar birleşmesi de SSB (single
strand binding) protein'lerin replikasyon çatalına yakın yerde iplikçiğe bağlanması ve bunun etkinliği ile
önlenir ve böylece ayrılan iplikçikler sentez için daima açık tutulurlar. Sentez,
birbirinden ayrılan parental iplikçiklerden 3' ¬ 5'
yönünde olanının karşısına, bu iplikçik kalıp olarak kullanılarak, DNA
polimerase enziminin katalitik etkinliği ile 5' ®3'
yönünde komplementer yeni bir iplikçik
çok çabuk olarak sentezlenir ve buna devam edilir (kesintisiz sentez). Bu yön (5' ® 3'),
DNA pol'ün aktivitesine de uygundur ve herhangi bir güçlük meydana gelmez. Ancak, DNA
pol enzimlerinin sentez yapabilmesi için bir primer
moleküle (başlangıç
basamağı) gereksinim vardır. Çünkü, DNA pol., gerek tek iplikçik DNA'da ve
gerekse primer olmayan DNA'da polimerizasyon yapamaz. Bu sorun, başlıca 3 tarzda
çözümlenebilmektedir. Bunlardan biri ve çok kullanılanı, parental DNA'nın
karşısına, 5' ® 3'
yönünde ve serbest 3'-OH ucuna sahip 8-14 bazdan oluşan primer RNA'ların
sentezleridir. Burası yeni DNA sentezi için başlangıç yeri oluşturur (veya, DNA'da
bir çentik açılarak serbest 3' -OH ucu
meydana getirilebilir veya RNA primerlerinin yerinde meydana gelen kısa protein primerleri de başlangıç oluşturabilirler).
Böylece bu başlangıç noktasından orijin alan ve parental iplik (3'¬ 5')
kalıp olarak kullanılarak, 3' -OH ucuna nukleotid trifosfatlar ilave edilerek
(trifosfatlardan biri bağlanır ve geri kalan iki fosfat molekülü serbest kalır), yeni
sentez 5'® 3'
yönünde diğer parental iplikçikle birlikte eş zamanlı olarak sürdürülür. Eğer,
yanlış bazlar sıraya girmişse, DNA pol. enzimi bunu hemen tanıyarak ekzonuklease
aktivitesi ile çıkarır yerine doğrusunu
koyar (düzeltme okuması). DNA sentezinin hızı, mRNA
sentezinden (transkripsiyon) 10 defa daha çabuktur.
Replikasyon çatalının ilerleme hızı dakikada 50.000 bp kadardır. Bu hız, S.
cerevisiae 'de 3600 bp/dk olarak
hesaplanmıştır. Parental
iplikçiğinin 5' ® 3'
yönünde olanında, ise, sentez kesintili olarak
sürdürülür ve kompleks bir karakter gösterir. Parental iplikçiğinin karşısına
sentezlenecek olan yeni iplikçik tersine 3' ¬ 5'
yönündedir. Bu istikamet, DNA polimerazın sentez aktivitesine terstir. Bu nedenle,
bazı güçlükler ortaya çıkmaktadır.
Bu
durum mikroorganizmalarda, şöyle çözümlenmektedir.Sentez, replikasyon çatalından
başlamakta ve ters yönde, parental iplikçik (5' ® 3')
boyunca kesintili olarak devam etmektedir. Diğer bir ifade ile, kesintili sentezin
yönü, replikasyon çatalının ilerleme yönüne terstir. Açılan parental iplikçiğe
SSB proteinleri bağlanarak çatalın devamlı açık tutulmasını ve sentezin devamlı
yapılmasını sağlarlar. Primase enzimi (dnaG) yaklaşık 10 bazdan oluşan primer RNA'ları sentezler ve bunun 3' -OH ucu
serbesttir. Bu uç basamak olarak kullanılarak yaklaşık 1000-2000 bazdan oluşan kısa
bir DNA segmenti (Okazaki fragment) sentezlenir.
Her Okazaki fragmenti ayrı bir primerden orijin alır. Bu sentezi, bir protein kompleksi
olan primosom başlatır. Primosomlar da
replikasyon çatalının ilerlediği yönde, diğer enzimlerle birlikte hareket ederler.
Ancak, bunun hareketi de Okazaki fragment'lerinin sentezi yönünün tersi istikametinde
olur. Okazaki fragmenti, bir önceki RNA primerlerine kadar uzanır ve burada durur.
Primerler DNA pol I'in etkisiyle çıkarıldıktan sonra bunların yerleri aynı enzimle
kapatılır ve iki DNA iplikçiği arası da DNA
ligase ile birleştirilerek iplikçik kesintili olarak tamamlanmış olur. Primase
enziminin çıkarılmasında da DNA pol III'ün rolü vardır. Böylece, 5' ® 3' yönündeki parental iplikçik kalıp olarak
kullanılarak, kesintili olarak devam eden ve sonunda tamamlanan 3' ¬ 5'
yönünde yeni bir DNA iplikçiği sentezlenmiş olur. Kesintili sentez işlemini de yine
DNA pol enzimi yapar. Replikasyon
çatalı ilerledikçe, sentez kesintisiz ve kesintili olarak aynı hızda ve sürede
tamamlanır. 4)
Terminasyon:
Sentezin sona ermesi de yine protein komplekslerinin aktivitesi sonu gerçekleşerek iki
iplikçik birbirinden tamamen ayrılır. 5)
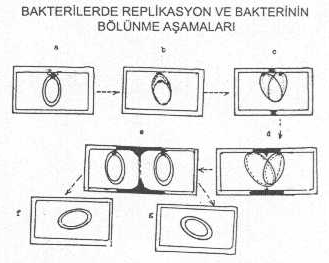
DNA'nın segregasyonu ve hücre bölünmesi: Genomik
DNA'nın sitoplasmik membrana bağlandığı yerde,
membranın sentezi nedeniyle giderek artan bir açılma meydana gelir. Bu açılma
ilerleyerek iki replike olan iplikçiği de birbirinden uzaklaştırır. Bunun sonunda,
bakterinin uzun ekseninde bir uzama gözlenir. Sonra, ortadan sirküler tarzda septum formasyonu meydana gelerek bakteri iki
hücreye ayrılır.
Kromozomun
iki hücreye ayrılmasında ve hücre divizyonunda oluşan aksaklıklar, bazı anormal
bakteri formlarının meydana gelmesine yol açar. Eğer septum formasyonu çok sık veya
yanlış yerden olursa kardeş hücrelerden birinde (küçük olanda) kromozom bulunmaz. Böyle hücrelere çekirdeksiz hücreler (anukleer hücreler) adı
verilir. Bazen de böyle hücreler kromozom ayrılmasının anormal olduğu durumlarda da
meydana gelir. Bunlarda da, hücrelerde kromozom yoktur. Ancak, septum formasyonu ve
boyutları normaldir. Septum
formasyonu inhibe olursa, kromozom normal olarak replikasyona devam eder. Ancak,
flamentöz formlar oluşur ve içinde birden fazla nukleus bulunur (multinukleer flamentöz formlar). Bazı mutantlarda (fts, flamentöz temperature sensitive), hücre
uzun flamentler oluşturur ve içinde çok sayıda nukleusa rastlanır. Buna karşın, par (partition) mutantlarında ise, kromozom
segregasyonunda bozukluk vardır. Çünkü, DNA anormal olarak hücrelere dağılır ve
bazı hücrelerde çekirdek bulunmaz. Defektler bu olguda DNA'da yerleşmiştir. Septum
formasyonuna neden olan gen, ftsZ'dir. Bu
gende oluşan mutasyon, septum formasyonunu önler ve flamentöz formların meydana
gelmesine yol açar. Mini hücre oluşumunda min B geninin rolü vardır. Bu lokusta meydana
gelen mutasyonlar minicell (bunlarda
genellikle çekirdeksizdirler) formasyonunu artırır. Kromozomun segregasyonunda da muk geni etkindir. 04.
Ökaryotiklerde Replikasyon Ökaryotiklere
ait replikasyon mekanizmasının incelenmesinde maya hücrelerinden fazlaca
yararlanılmıştır. Ökaryotiklerde
de prokaryotiklerde bulunan benzer enzim ve proteinler fonksiyoneldir (DNA pol., DNA ligase, topoisomerase, SSBP,
DNA'yı açıcı proteinler, vs.).
Ancak, bazıları yapısal farklılıklar gösterirler. Ökaryotiklerde 4 tür DNA
polimerase enzimi (a, b, d, e,) vardır, ayrıca bir de gama (g)
enzimi (mitokondrial) belirlenmiştir. DNA sentezi sırasında polimerase enzimi
fazlalaşmakta ve DNA sentezi hücrenin S-fazında meydana gelmektedir. Pol. b ve g enzimleri daha ziyade hücrenin istirahat halinde
iken bulunurlar. Pol. d, ayrıca, mitokondrial DNA'nın replikasyonunda etkilidir. Ökaryotiklerde
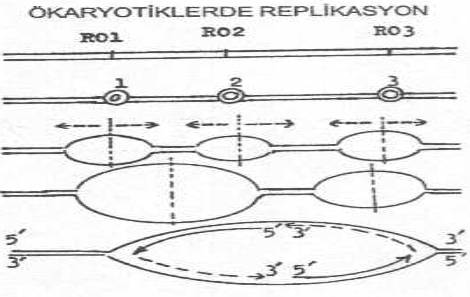
de replikasyon, prokaryotiklere benzemektedir. Ancak, kromozomun çok uzun olması
(bazıları bakterilerin bin katı) ve hücrenin S-fazında sentezin meydana gelmesi gibi
nedenler, prokaryotiklerden daha yavaş ve daha zaman alıcı bir DNA sentezine yol
açmaktadır. Belli bir süre içinde bu kadar fazla uzun kromozomun replikasyonu için,
sadece bir tane replikasyon orijini yeterli olamamakta ve bu nedenle birçok orijinden,
aynı anda veya bazen değişik zaman aralıklarında başlayan, bidireksiyonal replikasyon meydana gelmektedir.
Replikasyon, sağa ve sola doğru genişleyen orijinlerin birbirine değdiği yerde son
bulur.
Ökaryotiklerin
kromozomları histonla birleşmiştir (nukleosome) ve
replikasyon sırasında histondan ayrılırlar. Replikasyonda,
DNA sentezi her iki iplikçikte aynen bakterilerde olduğu gibi sürdürülür (biri
kesintisiz ve diğeri kesintili). Maya
hücrelerinde yaklaşık 400'e yakın orijin (ARS,
autonomously replicating sequence) bulunmaktadır. 05. Diğer Replikasyon Modelleri 05.01. Mitokondrial DNA (mt DNA)
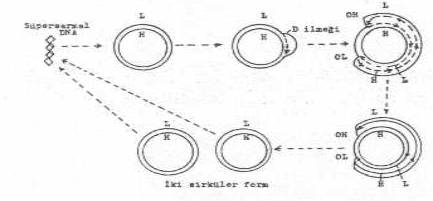
Replikasyonu Memeli
hücrelerinin mitokondrisi yaklaşık 5 µm uzunlukta (MA: 10x106, 15 000 bp) olup sirküler, süper sarmal bir
DNA yapı özelliği gösterir. mtDNA'nın replikasyonunda, DNA pol, d
enziminin aktif rolü vardır. Fare
L-hücreleri mitokondrisi üzerinde yapılan çalışmalarda, mtDNA'nın sentezine her iki
iplikçiktede diğer mekanizmaların aksine, kesintisiz olarak devam ettiği ortaya
konulmuştur. Ancak, replikasyonda bazı farklı yanlar da bulunmaktadır. Şöyle ki, biri ağır (H) ve diğeri de hafif (L) iplikçiklerde iki replikasyon orijini
(OH ve OL) yer almaktadır. DNA'da, önce, H-iplikçiğindeki orijinden (OH) başlayarak
bir D-ilmeği meydana gelir ve ilmek
genişleyerek unidireksiyonal olarak sentez devam eder. H-sentezi, 2/3 uzunluğa
erişince, bu defa L-iplikçiğindeki OL bölgesinde sentez başlar ve H-ye ters olarak bu
da unidireksiyonal
olarak ve kesintisiz devam eder. Sonra iki sirküler DNA birbirinden
ayrılır, sentez tamamlanır ve DNA iplikçiğinde 100'e yakın super sarmal meydana
gelir.
05.02. Rolling Circle Replication Bu
tür replikasyon modeline bazı fajlarda (ØX174,
Lambda) ve F-faktörünü kromozomlarında
taşıyan Hfr-hücrelerde
rastlanmaktadır. 1)
Lambda (l)
fajında replikasyon: Bu faj
konakçı bakteri içinde başlıca iki formda bulunur. Biri konakçıyı lize eden (litik
form) ve diğeri de konakçının
kromozomunun belli bir bölgesine (biotin geni
ile galaktoz geni arasına) yerleşerek
herhangi bir bozukluk oluşturmayan latent form.
Böyle formu oluşturan lambda fajına profaj adı verilir. Litik
formda faj ürerken, oluşan bazı sirküler formlarında bir iplikçikte meydana gelen
kopma nedeniyle 5' -ucu dışa doğru açılarak uzanır. Buna karşın, 3' -ucu ise
ortadaki sirküler tek iplikçiğin etrafında yeni komplementer iplikçik sentezler. Ayrılan 5'
-ucu, normal boyutuna veya bunun 2-3 katı uzunluğa ulaştığında paketleme enzimi Cos bölgelerinden iplikçiği
keserek normal boyutlarda lineer çift iplikçik DNA segmentleri meydana getirir. Bunlar
paketleme enzimleri yardımıyla, hücre içinde sentezlenen boş faj başlığı içine girerler ve böylece olgun
infektif fajlar meydana gelir. 2)
Øx174 fajında replikasyon: Tek
iplikçik DNA fajında da rolling circle
replikasyon modeli sentez
görülmektedir. Faj
DNA'sı sirküler tek iplikçik ve (+) polaritelidir. Hücre içinde buna (–) polariteli
bir iplikçik daha sentezlenerek çift iplikli hale getirilerek replikatif formlar (RF) oluşturulur. Bu
formlar, genellikle, super sarmal bir
yapıya sahiptirler. Topoisomerase enzimi süper heliksi açarak istirahat haline getirir.
Protein A, (+) polariteli iplikçiğe bağlanarak bir çentik açar ve protein A, 5'
-ucuna bağlı olarak iplikçiği ayırır. Diğer 3' -ucu ise sentezi devam ettirir.
Ayrılan 5' -ucunun bulunduğu DNA iplikçiğine stabilizan tek iplikçik proteinler (SSB)
bağlanarak bunun (–) polariteli iplikçikle tekrar birleşmesi önlenir. Rep
proteinleri de iki iplikçik arasındaki bağları çözerek, DNA' iplikçiklerini
birbirinden ayırmaya devam eder. Sentez 3' -ucundan, (–) polariteli iplikçik
etrafında ilerler. Ayrılan 5' -ucu tam bir dönüş uzunluğuna ulaşınca serbest
uçtaki protein A bu defa (–) iplikçik etrafında yeni sentezlenen (+) iplikçiğe
bağlanarak tekrar çentik açar ve replikasyon böylece devam eder. Protein A'dan
kurtulan (+) iplikçik sirküler forma dönüştürülerek faj başlığı içine
paketlenir. Bu iki
fajın replikasyonları hakkında fajlar bahsinde
gerekli bilgiler verilmektedir. 3)
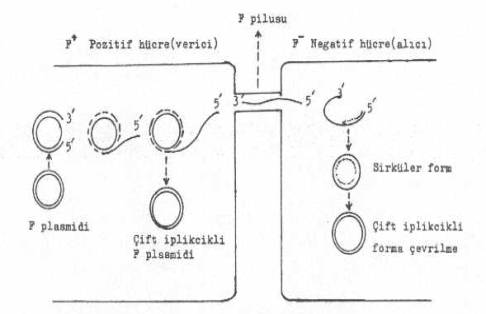
Hfr hücrelerde replikasyon: E.
coli
'lerde
bulunan ve seks pilusunun formasyonuna neden olan seks faktörü (F-faktörü) bir plasmiddir. Bu
faktör E. coli içinde ya sitoplazmada
bulunur ve E. coli 'yi F+
hale getirir veya bakterinin genomu ile birleşerek Hfr (High frequency recombination) oluşturur. Bu
son durumda bakteri, kendi kromozomunu ve F-faktörünü transfer için seks pilusu
sentezler. Bu pilus, kendisinde F-faktörü bulunmayan (F¯) hücreye genetik materyalin
aktarılması için bağlanarak bir köprü görevi yapar. Genetik materyal, oluşan bu
konjugasyon köprüsünden geçerek (F¯) hücreye transfer edilir. F
plasmidinde, transfer için yaklaşık, 35 kb'lık bir bölge (transfer bölgesi) etkilidir. Pilus formasyonu
için de 12 gene (tra gen) ihtiyaç
vardır. F+ bir hücre, diğer F+ bir hücre ile pilus aracılığı
ile temasa gelemez. Çünkü, tra S ve tra T genleri bakteri yüzey eksklusiyon proteinlerinin sentezine neden
olur ve bunlar pilusla sağlanan teması önlerler. Böylece F+ veya Hfr bir
hücre ancak, F¯ hücre ile ilişki kurabilir.
F-faktörünün
transferi oriT (origin
transferi) bölgesinden başlar. Burası, transfer orijininin bir ucunda lokalize
olmuştur. OriT'de, Tra Y ve/veya
Tra I proteinleri
tarafından bir çentik oluşturulur. Proteinler, sonra, DNA'ya bağlanır ve yaklaşık
200 bp uzunlukta DNA açılır. TraY/TraI multimeri 5' -ucundan hareket ederek bir
saniyede 1200 bp açar ve açmaya devam eder. Sonra, 5' -ucu tekrar alıcı bakteriye
yönelir. Konjugasyon köprüsünden alıcıya geçince bu tek iplikçiğe ikinci
iplikçik (komplementer) sentezlenerek sirküler bir duruma gelir. 05.03. Plasmidlerde Replikasyon Bakterilerde
sitoplazmada serbest olarak bulunan plasmidler replikasyon orijinine sahip olduklarından,
bakteri genomuna bağlı olmaksızın, bakteri hücresi içinde otonom olarak kendi
replikasyonunu yönetebilirler. Replikasyon tarzları, bakteri kromozomuna benzerdir.
Ancak, plasmidler bakterinin kromozomu ile birleşince (episom), aynen profajlar gibi, DNA'nın bir
parçası gibi olurlar, onunla birlikte replike olurlar ve kardeş hücrelere transfer
edilirler. 05.04. Replikasyon İnhibitörleri DNA
replikasyonu, sentez sırasında, çeşitli aşamalarda bazı kimyasal maddeler
tarafından inhibe edilirler. Bu inhibitörler çok değişik karakter gösterdikleri gibi
etkinlikleri de oldukça farklıdır. Bazıları nukleotid prekürsör sentezini inhibe
eder, bir kısmı nukleotid analoğu olarak DNA'ya inkorpore olur ve bazıları da DNA'ya
bağlanarak aktivitesine mani olur. Enzimlere, özellikle, DNA polimerase ve replikasyonda
fonksiyonu olan diğer enzimlere bağlanarak görevini aksatan ve bozan maddeler de
vardır. Bazı
fiziksel ajanlar (UV-ışınları, X-ışınları ve gama ışınları) DNA'ya zarar verir
ve replikasyonda bozukluklar meydana getirir. Böyle direkt etki yapan ajanların
dışında, bazı kimyasal maddeler de sentez inhibitörü olarak indirekt etkiye
sahiptirler. Örn. Colchicine ve diğer
mikrotubuler parçalayıcıları (Vinca
alkaloidleri, vincristine ve vinblastin gibi). Replikasyon
inhibitörleri, etkiledikleri bölgelere göre 3 gruba ayrılmaktadırlar. a)Nukleotid
biyosentezini inhibe edenler (pürin, pirimidin, folate, deoksiribonukleotid sentez
inhibitörleri vb.). [1]
Kaynak: Temel
Mikrobiyoloji
|
||||||||||||||||