|
Bakterilerin Genetik Karakterleri Prof. Dr. Mustafa Arda
Ankara Üniversitesi
Veteriner Fakültesi 01. Genetik Materyallerin Yapısı
01. Genetik Materyallerin
Yapısı
Nukleik
asitler, prokaryotik ve ökaryotiklerin, virus, plasmid, faj, transpozon,
IS-elementlerinin temel makromolekülünü oluşturduğundan, ayrıca bütün yaşamsal
fonksiyonları ve kalıtsal özellikleri de yönettiğinden çok önemli rollere
sahiptirler. Bu bölümde daha ziyade prokaryotiklerin (bakteri), virus, plasmid ve faj
genomlarının moleküler yapıları ve işlevleri hakkında genel mikrobiyolojiye
dayalı, özlü ve temel bilgiler verilecektir. Nukleik
asitler, ilk defa, İsviçreli bir fizyolog olan Friedrich Mischer tarafından 1869
yılında bildirilmiştir. İrin içindeki beyaz kan hücrelerinde proteinlerden ayrı
olarak asit karakterde bazı maddelerin de varlığını ortaya koyan araştırıcı,
bunlara nuklein adını vermiştir. Çalışmalarını sonraları salmon balıklarının
spermatozoidlerinin çekirdekleri üzerinde yoğunlaştıran F. Mischer, bu
araştırmalarını 1874 yılında yayımlamıştır. Benzer konularda incelemelerini
sürdüren diğer bir bilim adamı da, Richard Altman (1889), asit karakterleri nedeniyle,
nukleinleri nukleik asit olarak yeniden isimlendirmiştir. Alman kimyacı, Robert Feulgen,
nukleik asitleri, kendi adı ile anılan spesifik bir boyama yöntemi (Feulgen) ile
boyamayı başarmıştır (1920). Aynı tarihte, Rockefeller Enstitüsü'nde, Phoebus
Levine, nukleik asitlerin yapısında pentoz şekerinin bulunduğunu açıklamıştır.
Oswald Avery (1930), nukleik asitlerin (DNA) genetik bir madde olduğunu bildirmiştir. Watson
ve Crick (1953), DNA'nın çift iplikcikli ve sarmal bir yapıya (double-helix) sahip
olduğunu belirtmişlerdir. Bu tarihten sonra, gerek prokaryotik ve gerekse ökaryotik DNA
üzerinde çok yoğun çalışmalar yapılmaya başlanmış ve halen de aynı hızda devam
etmektedir. Bu araştırmalar sayesinde DNA'nın fonksiyonları hakkında çok
ayrıntılı ve değerli bilgiler elde edilmiştir. Ayrıca, genetik materyal olarak, RNA
ve mRNA üzerinde de benzer tarzda incelemeler yapılmıştır. Hücrelerdeki
bütün biyolojik olayları (fizyolojik, biyokimyasal, genetik vs.) yöneten, genetik
bilgiler taşıyarak bunların nesillere aktarılmasında önemli fonksiyonları bulunan
nukleik asitler (DNA ve RNA), hücrelerde bir merkezi jeneratör görevine sahip
bulunmaktadır. Bu makromolekül, başlıca iki yapısal özellik taşımaktadır. 1)
Deoksiribonukleik asit (DNA) 02. Prokaryotik Deoksiribonukleik
Asit (DNA) Bakterilerde
DNA, hücre kuru ağırlığının yaklaşık %2-3 kadarını oluşturmakta ve
mikroorganizma türüne göre değişmek üzere de 0.01 pg (1 pg= 10-12 g) ağırlığında olabilmektedir.
Mikoplasmalarda DNA'nın molekül ağırlığı (MA) 4x108 dalton, hemofiluslarda MA: 8x108 dalton, B. subtilis 'de 3.9x109 ve E.
coli 'de 2.6x109 kadardır (1
dalton= 1.67x10-24 g). DNA'nın
uzunluğu, E. coli 'de yaklaşık 1.1-1.5 mm
kadar olup 4x106 nukleotid çiftinden
(baz çifti) oluşmaktadır. Bu uzunluk E. coli
'nin yaklaşık 500 katı kadardır. Nukleotid
birimlerinden (monomer) oluşan ve bir polimer olan DNA'nın yapısında 3 temel komponent
bulunmaktadır. Bunlar da; 1)
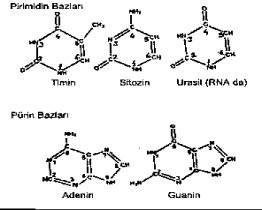
Pirimidin ve pürin bazları, 02.01. Pirimidin Bazları DNA'nın
yapısında yer alan pirimidin bazları, 4 karbon ve 2 nitrojen atomundan oluşmuş tek
halkalı (6 köşeli) bir temel yapı özelliği gösterirler. Bu çatının serbest
uçlarına bazı atom veya atom grupları bağlanarak pirimidinler meydana gelmişlerdir.
Nukleik asitlerde başlıca iki tür pirimidin bazı bulunmaktadır. Bunlar da, Sitozin
(C: 4 amino-2-oksipirimidin) ve Timin (T: (2,4-dioksi 5-metil pirimidin). Primidin
bazlarında oksijen, keto formunda (C=0) bulunur.
Pirimidin
bazlarından olan Urasil (U: 2,4-dioksi pirimidin) timin yerine RNA'nın yapısında
bulunur. 02.02. Pürin Bazları Bir
pirimidin derivatı olarak kabul edilen pürin bazlarında, biri primidin (6 atomlu) ve
diğeri de buna birleşmiş 5 atomlu imidazol halkasından (3 karbon + 2 nitrojen atomu)
(meydana gelmiş) iki halka bulunur. DNA'da
iki tür pürin bazı bulunmaktadır. Bunlardan biri, Adenin (A: 6-amino pürin) ve
diğeri de Guanin (G: 2 amino; 6 oksipürin)'dir. Pirimidin
ve pürin bazları hidrofobik bir karaktere ve planer bir yapıya sahiptirler. Bazı
mikroorganizmalarda (fajlarda) nukleik asitlerin yapısında, yukarıda belirtilen temel
bazların yanı sıra minor bazlar olarak adlandırılan,
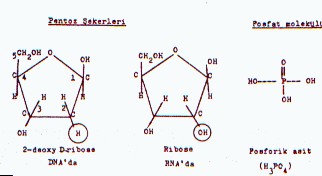
5-metil sitozin (5'-MC) ve 5'-hidroksi metil urasil (5'-HMU) bulunmaktadır. 02.03. Pentoz Şekeri DNA'nın yapısındaki ikinci önemli komponenti, 5 karbonlu bir pentoz olan D-riboz (C5H10O5) oluşturur. Ancak, bu şekerin 2. pozisyonunda bulunan karbon atomuna bağlı bir oksijeni
bulunmamaktadır (C5H10O4).
Bu nedenle de şeker, 2-deoksi-D-riboz ve nukleik asit de, buna bağlı olarak,
deoksiribonukleik asit olarak, adlandırılmaktadır.
02.04. Fosfat Grubu Nukleik
asitlerin (DNA ve RNA) yapısında bulunan 3. komponent de fosfat molekülüdür (H3PO4).
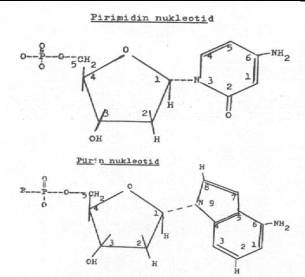
Fosfat, hem DNA'da ve hem de RNA'da aynı yapıdadır. 02.05. Nukleosidler Nukleosidler:
Pirimidin (T,C) veya pürin bazlarından (A,G) birisi ile pentoz şekeri
(2-deoksi-D-riboz) birleşerek nukleosidleri meydana getirirler. Her iki molekül
arasındaki bağlanma, pentoz şekerinin 1. pozisyonunda bulunan karbon atomunun,
pirimidinlerin 3'-pozisyonundaki N-atomu ile ya da pürinlerin 9'-pozisyonundaki N-atomu
ile beta glikozid bağları yardımı ile gerçekleştirilir. Nukleosidler
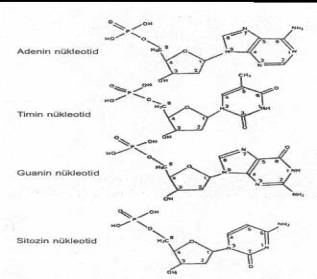
de, orijinlerine göre, pirimidin nukleosidler (timin deoksiribonukleosid ve sitozin
deoksiribonukleosid) ve pürin nukleosidler (adenin deoksiribonukleosid ve guanin
deoksiribonukleosid) olmak üzere iki kısıma ayrılmaktadırlar. 02.06. Nukleotidler Nukleosidler
de fosfat molekülleri ile birleşerek nukleotidleri oluştururlar (deoksiribonukleotid).
Bu birleşme, pentoz şekerinin 5'-pozisyonundaki karbon atomu ile fosfat molekülünün
oksijeni arasında kurulan ester bağı ile sağlanır. Nukleotidler de yapısındaki
bazların türlerine göre pirimidin nukleotidler (timin deoksiribonukleotid ve sitozin
deoksiribonukleotid) ve pürin nukleotidler (adenin deoksiribonukleotid ve guanin
deoksiribonukleotid) olarak iki kısıma ayrılırlar.
02.07. Polinukleotidler Yan
yana bulunan nukleotidler (monomer), birbirleri ile fosfodiester bağları ile birleşerek
polinukleotid zincirlerini (polimer) oluştururlar. Fosfat
grubu, bitişik nukleotidin pentoz şekerinin 5' pozisyonundaki karbon atomu ile diğer
komşu şekerin 3'-pozisyonundaki karbon atomu arasında fosfodiester (O-P-O) bağı
kurar. Bağlantının yönü 5'-P terminusundan 3'-OH terminusuna doğru olduğundan
böyle bağlantıya 5'®3'
fosfodiester bağı adı verilir. DNA'nın sentezi sırasında nukleotid trifosfatlar
pentoz şekerinin 3'-OH ucuna bağlandığından, polinukleotidin büyümesi bu uçtan
başlayarak 5®3'
yönüne doğru devam eder .
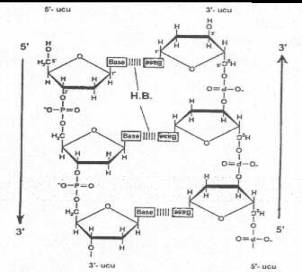
Bir
bakteri genomunda bu tarzda büyüyen, birbirine karşılıklı hidrojen bağları ile
birleşmiş iki adet polinukleotid iplikçiği vardır. Bu iki iplikçikten, birinin 5'-P ucu serbestse, bunun karşısında bulunan
diğer iplikçiğin 3'-OH ucu serbesttir. Bu durum, her iki iplikçiğin diğer uçları
için de benzeridir. Böylece iki iplikçik bir polariteye sahip olur ve iplikçikler
birbirine anti-paralel bir durum gösterirler. Buna göre, iplikçiklerden birinin
büyüme yönü 5'®3'
ise, diğerinin büyüme yönü tersine 3'¬
5' olmaktadır. Bu
tarz bir yapı özelliği gösteren DNA'da, bir iplikçikte bulunan pirimidin bazları
(timin, sitozin) diğer iplikçikte bulunan pürin bazları (A, G) ile karşılıklı
hidrojen bağları ile birleşmişlerdir. Adenin timinle 2 hidrojen bağı (A= = = T) ve
guanin de sitozinle 3 hidrojen bağı (Gº
º
ºC)
ile birleşerek orta eksende yer alırken şeker-fosfat omurgası da dış tarafta
bulunur. G ile C arasında 3 hidrojen bağının bulunması nedeniyle, A ile T
bağlantısından daha sağlam bir birleşme oluşturur. Karşılıklı birleşmiş olan
nukleotidler baz çiftlerini meydana getirirler ve böylece DNA çift iplikcikli, sarmal
ve sirküler bir yapı karakteri gösterir. İki
nukleotid iplikçiği, orta eksen etrafında sarmal sirküler (kapalı) ve çift
iplikcikli bir yapı karakteri gösteren tek bir moleküldür. Bu strüktür üzerinde
biri büyük ve diğeri de küçük olmak üzere iki oluk meydana gelir.
Bir
iplikçikte bulunan pirimidin bazları, karşı iplikçikteki pürin bazları ile
birleştiğine göre, DNA'da ne kadar pirimidin varsa o kadar da pürin bazı olacak
demektir. Diğer bir ifade ile, sayısal olarak A = T ve G= C olur. Aynı zamanda, A + G =
T + C ve A/T = 1 ve G/C = 1'dir. Ancak, A + T/G + C oranı değişik bir durum gösterir.
Yüksek hayvan ve bitkilerde bu oranın değeri büyük olmasına karşın, bazı bakteri
ve viruslarda büyük veya küçük olabilmektedir. Örneğin C. perfringens 'de A + T/G + C = 70/30 ve A. bovis 'de ise 26/74'dür. Dört
temel bazın (A,T,G,C) değişik sırada yan yana ve karşılıklı birleşmesinden
oluşan DNA iplikçikleri hücreler ve canlılar için çok önemli olan genetik bilgileri
taşırlar ve bu bilgiler bazların diziliş sırası ile yakından ilgilidirler. Orta
eksende yer alan bazlar bu bilgilerin korunmasını güvence altına almış olurlar.
Aynı türün DNA kompozisyonu (diziliş sırası ve sayısı) birbirinin aynıdır,
sabittir ve değişmez. Farklı türlerin DNA'ları birbirine uymaz. Bu durum türler
arası genetik farklılıkları oluşturur. Genetik düzeyde oluşan değişmeler
(mutasyonlar), yeni nesillerde farklı fenotipik karakterde bireylerin meydana gelmesine
neden olurlar (mutant). Ancak, normal koşullar altında böyle olgular çok nadirdir (10-7-10-10
gibi). Bir
iplikçikte bulunan baz türü ve sırası karşı iplikçikteki baz türünü ve
sırasını da belirler ve böylece birbirini bütünler ve tamamlar (komplementer) bir
karakter gösterirler. Bu duruma göre, bir iplikçikteki baz türü ve sırası: AT G C C
C... A ise, karşı iplikçikleri T A C G G T .....T bazları bulunması gerekecektir. DNA'nın
sarmal ve özellikle, çift iplikcikli olmasının önemi çok fazladır. Çift iplikcikli
olmakla genetik bilgiler garanti altına alınır ve güvenceleri sağlanır. Ayrıca, DNA
kendine bir replikasyon mekanizması kurarak, bir iplikçik diğerinin sentezi için
kalıp ödevi görebilecek duruma gelir. Bir iplikçikte oluşan kopma ve zedelenmeler,
sağlam olan karşı iplikçik kalıp olarak kullanılarak, kopan yerdeki bazların yerine
polimeraz enzimi tarafından yenileri yerleştirilir. İplikçiklerin her biri aynı
genetik bilgilere sahip olduğundan transkripsiyonda sadece bir iplikçik görev alır ve
bu da, 3¬
5' yönünde olanıdır. Baz çiftleri, aynı
zamanda, replikasyonun doğru yönde ilerlemesine de yardımcı olurlar. Polinukleotid
iplikçiklerinde bulunan 4 bazın (A,T,G,C) diziliş sırasında çok önemli bilgilerin
şifreleri (genetik kodlar) saklı olarak bulunur. Bu bazlardan yan yana bulunan 3 tanesi
bir amino asitin kodunu oluşturur (triplet veya kodon). Bu üçlü sistemde 64 (43) kodon bulunmaktadır. Bu duruma göre, 20
amino asit için 64 kodon vardır ve her bir amino asit için de en az bir veya birden
fazla kodon bulunacak demektir. Örneğin, fenil alanin için UUU ve UUC, serin için UCU
ve UCC veya valin için GUU, GUC, GUA ve GUG gibi birden fazla triplet fonksiyon yapar.
Eğer, her amino asit için 2 bazdan oluşan bir kodlama olsaydı, 20 amino asit için 16
(42) kodon bulunacaktır ve bu sayı
da yetersiz kalacaktır. Böylece, 3 baz olmakla veya kodon sayısı artmakla her amino
asit için hem seçenek ve hem de spesifite arttırılmış oluyor. Kodonlar arasında
UAA, UAG ve UGA gibileri hiçbir amino asitin şifresi olmayıp bunlar translasyonda,
sentezlenen peptidin bitişinin sinyalini veren kodonlardır. Bir
bakteride genetik kodlar ve özellikleri şöyledir. 1)
Kodonlar triplettir: Genetik bilgileri saklayan kodlar, 3 bazın birleşmesinden meydana
gelmişlerdir. 2)
Kodonlar değişkendir: Her amino asit için bir veya birden fazla triplet bulunmaktadır.
Örn., fenilalanin için UUU ve UUC, serin için UCU ve UCC, vs. birden fazla kodon
bulunur. Bu örnekleri daha da çoğaltmak mümkündür. 3)
Kodonlar birbirlerine çakışmazlar: Bir triplette bulunan bazlar, aynı anda, başka bir
amino asitin kodonu olamaz. Diğer bir ifade ile aynı harflerle oluşan bir kodon, ancak
bir amino asitin şifresini teşkil eder. Ayrıca, yan yana bulunan bazlardan biri, bir
önceki veya sonraki amino asitin kodonunun bir harfini (bazını) oluşturamaz. Bu
nedenle, kodonlar birbirlerinden tam bağımsızdırlar. 4)
Kodonlar arasında boşluk yoktur: Prokaryotik mikroorganizmalarda (bakterilerde) kodonlar
DNA veya mRNA üzerinde yan yana aralıksız olarak devam ederler. İki triplet arasında
herhangi bir baz(lar) veya sekans bulunmaz ve bu nedenle arada intron (kodlamayan sekans)
yoktur. Bu durum, genler için de geçerlidir. Buna karşın, viruslarda ve
ökaryotiklerde, birbirleriyle ilişkili genler olsa bile, bazen aralarında 2000 veya
daha fazla bazlık kodlamayan sekanslar (intron) bulunmaktadır. Ancak, DNA'dan mRNA
sentezlenirken bu intronlar primer mRNA' ya aktarılmasına karşın, olgun mRNA'nın
meydana gelmesinde intronlar çıkarılır ve sadece kodlayan genler (eksonlar) yan yana
getirilerek birleştirilir ve böylece translasyon için sadece olgun mRNA kullanılır. 5)
Kodonlar üniversaldir: Bir triplet, bakteride hangi amino asitin şifresi ise,
ökaryotiklerde ve diğer organizmalara ait DNA'lar da aynı amino asitin kodonudur. Bu
durum da, canlıların genetik materyallerinin amino asit şifrelemesi yönünden
birbirlerinin benzeri olduğunu göstermektedir. Ayrıca, baz türlerinin de hemen hemen
aynı olduğunu anlamak mümkündür. Bakterilerde
DNA, iki adet polinukleotid iplikçiğinin sarmal bir tarzda karşılıklı
birleşmesinden oluşmuş ve sirküler bir yapı karakteri gösterir (double heliks, çift
sarmal). Aynı zamanda, bu polinukleotid iplikçiğinin iki ucu da kapalıdır (sirküler)
ve tek bir molekül halindedir. Uzunluğu, bakterilerin türlerine göre değişmek
üzere, bakteri boyunun 400-500 katı kadardır (1.1-1.5 mm). Ancak, B. burgdorferi 'de lineer bir kromozomun olduğu
bildirilmiştir. 02.08. Başlıca DNA Tipleri
Mikroorganizmalarda
bir çok tipte DNA bulunabileceği belirtilmiştir. Ancak, bunların çoğu deneysel
koşullarda rastlanan tiplerdir (A, B, C, D, E ve Z gibi). Bu formların saptanmasına
temel teşkil eden esaslar kısaca şöyledir. 1)
Heliksin bir tam dönümüne isabet eden baz sayıları, Yukarıdaki
kriterlerin ışığı altında, prokaryotiklerde (bakterilerde) rastlanan en önemli DNA
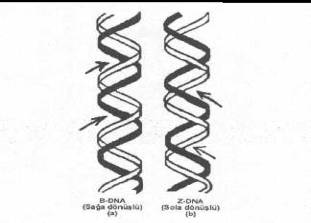
tipleri 3 tanedir. Bunlar da, B-DNA
tipi: Watson-Crick modeli DNA, bu tür için en iyi bir örnektir. Bu tür doğal DNA'nın
şeker-fosfat omurgası düzgün ve sağa dönüşlüdür. Sarmal dubleksin çapı 2.0 nm,
her bir 3.4 nm'de tam bir dönüş yapar ve her bir dönüşe 10 nukleotid çifti isabet
eder. Her bir baz çiftinin kalınlığı 0.34 nm olup bazlar orta eksene dikeydir ve
bazlar arasında (birbirine bitişik nukleotidler arasında) orta eksene göre 36°’lik
bir açı vardır. A-DNA
tipi: Deneysel koşullarda elde edilen bu tip DNA, süspansiyon halinde iken suyun
çıkarılması (dehidrasyon) durumunda ve %70-75 etanol içinde meydana geldiği
açıklanmıştır. Böyle ortamlarda DNA'da büzülmeler oluşmakta ve her bir tam
dönüşe 11 baz çifti isabet etmektedir. Sağa dönüşlü bir omurgası bulunan
A-DNA'da bazlarının kalınlığı 0.26 nm, orta eksene göre bazlar arasında 33°'lik
açı bulunmakta ve tam bir dönüş 2.8 nm
uzunluktadır. Heliksin çapı 2.3 nm kadardır.
Z-DNA
tipi: Bu DNA tipi de çift iplikcikli ve sarmal bir yapı özelliği taşımasına
karşın omurgası zigzaglı ve sola dönüşlüdür. Bu tür DNA'da bir tam dönüşe 12
baz çifti (6 dimer) isabet eder. Bazların kalınlığı 0.37 nm ve heliksin
kalınlığı ise 1.8 nm kadardır. Tam bir dönüş 4.5 nm olup her dimer arasında
-60°lik açı vardır. 02.09. Diğer Bazı Yapısal Özellikleri Nukleik
asitlerin, proteinlerde de belirlendiği gibi, bazı yapısal özellikleri bulunmaktadır. 1)
Nukleik asitlerin primer yapıları: Nukleik asitlerin komponentlerinin birbirlerine
bağlanma özellikleri (anti paralellik, 5'®3'
ve 3'®5'
yöndeki bağlantıları, 5'-ucunda P ve 3'-ucunda OH molekülünün bulunması, bazların
genetik informasyon taşımaları, 4 bazdan oluşma durumu, vs) 2)
Nukleik asitlerin sekonder yapıları: Bu yapı içinde, nukleik asitlerin sarmal
yapıları, her sarmala isabet eden baz sayıları, karşılıklı bazlar arası hidrojen
bağları, DNA'nın yapısı (A, B, Z DNA formları), vs bulunmaktadır. 3)
Nukleik asitlerin tersiyer yapıları: Bu kavram içinde DNA'nın sirküler, lineer ve
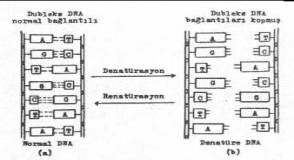
süpersarmal formları yer almaktadır. 02.10. Denatürasyon ve Renatürasyon Bakterilerden
saf olarak elde edilen DNA süspansiyonu 100°C 'ye kadar yavaş yavaş ısıtılırsa
(veya kimyasal yöntemlerle) bazlar arasındaki karşılıklı hidrojen bağlarında
çözülmeler meydana gelerek iki iplikçik birbirlerinden ayrılabilir (denatürasyon).
Eğer bu süspansiyon, yavaş yavaş soğutulursa bu defa da karşılıklı hidrojen
bağları tekrar kurulur ve molekül eski formunu alır (renatürasyon). DNA'nın bu
özelliğinden yararlanılarak suni veya melez (hibrid) DNA molekülleri oluşturmak
(hibridizasyon) ve bunları çeşitli araştırmalarda, mikroorganizmaları sınıflamada,
gen transferlerinde veya diğer genetik manipulasyonlarda kullanmak olanak dahiline
girmiştir. Hibridizasyon denemelerinde çok yararlanılan böyle çalışmalarda, hibrid
moleküller oluşturmak ve sonra da bunları işaretli problarla saptamak mümkündür.
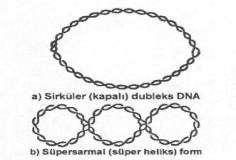
Prokaryotiklerin
(bakteriler) DNA'ları başlıca 2 topolojik formda bulunmaktadır. a)
Sirküler dubleks DNA formu: Bakteriler, bazı fajlar (PM2, vs.), iplikcikli sarmal
sirküler bir karaktere sahiptir. b)
Süpersarmal (süperheliks) DNA formu: Fajlarda, plasmid ve viruslarda daha fazla
rastlanılan bu form, dubleks sarmal DNA'nın serbest ortamda kendi ekseni etrafında
360° ve birkaç kez dönmesinden kaynaklanmaktadır.
Süpersarmal DNA formuna daha ziyade sirküler sarmal DNA' larda fazlaca rastlanılmaktadır. Süperheliks formları önceleri küçük DNA moleküllerinin (virus, faj, plasmid, mitokondrial DNA, vs) özel bir karakteri olarak düşünülmüştür. Bazı araştırmacılar da, DNA izolasyonları sırasında oluşan artifaktlar olarak kabul etmişlerdir. Bugün ise, bu formların prokaryotik hücrelerin (ve ayrıca faj, virus, plasmid) DNA'larının önemli bir karakteri olduğu, replikasyon, transkripsiyon ve rekombinasyonlarda fonksiyonlarının bulunduğu anlaşılmıştır.
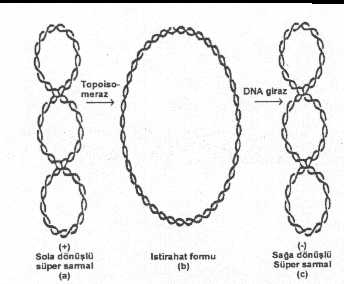
Süpersarmal
DNA, ya sola dönüşlü pozitif süpersarmal veya sağa dönüşlü negatif süpersarmal
formda bulunabilir. İn vivo koşullarda ise daha ziyade negatif süpersarmala
rastlanılmakta ve DNA üzerine de torsiyonal bir stres yaratarak açılmasını
kolaylaştırmaktadır. Buna karşın, pozitif süpersarmal daha sağlam ve sıkı
yapısal bir özellik taşımaktadır. Bir DNA'da, çok sayıda sarmal oluşabilir. Aynı
zamanda, süpersarmal form, DNA'nın santrifüjde daha çabuk çökmesini de sağlar. Hücrelerde
süpersarmalın varlığı bazı tekniklerle ortaya konulabilir. 1)
Elektron mikroskopla direkt muayene: Hücrelerden elde edilen DNA'nın (virus, faj,
plasmid, vs.) elektron mikroskopla direkt muayenesi ile süpersarmal durumu kolayca
görülebilir. Bu tarzda fotoğrafları çekilmiş bir çok DNA'nın süpersarmal
formları gözlenmiştir. 2)
Sedimentasyon metodu: Gerek pozitif ve gerekse negatif süpersarmal DNA'lar sirküler ve
sarmalsız olanlardan daha çabuk çökme özelliğine sahiptirler. Bu nedenle de
süpersarmal DNA'lar santrifüj tüpünün dibinde ve sadece sirküler olanları da bunun
üstünde bulunurlar. 3)
Elektroforezis yöntemi: İzole edilen DNA'lar agarose jele konarak elektroforezise tabi
tutulursa, süpersarmal DNA'lar ile istirahat halinde olanlar veya süpersarmal
içermeyenler birbirlerinden kolayca ayırt edilebilirler. Süpersarmal DNA'lar daha
ileride bir lokalizasyona sahip olmalarına karşın, diğerleri başlangıç noktasına
yakın bulunurlar. DNA
molekülünün negatif veya pozitif süpersarmal bir duruma gelmesinde ve bu
sarmalarının açılarak istirahat formunu almasında Topoisomeraz olarak adlandırılan
enzimlerin rolü bulunmaktadır. Bu enzimler de başlıca iki tane olarak saptanmıştır. Topoisomeraz-I: Yaklaşık 100000 molekül ağırlığında olan ve monomerik bir yapıya sahip olan bu enzim (Topo-I) negatif süpersarmal DNA molekülünün tek bir iplikçiğinde ve geçici bir süre için kopmalar ve tekrar birleşmeler yapar. Enzim, süpersarmal DNA'yı açarak istirahat formuna dönüştürür. Bu fonksiyonu için enerjiye gereksinim duyulmaz. Enzim, DNA iplikçiklerinden birine bağlanarak kopma (veya açılma) yapar ve ikinci sağlam DNA iplikçiğinin bu açıklıktan geçmesini sağlar. Sonra, ayrılan iki uç enzimle hemen kapatılır. Bu açılma
ve kapanma, DNA molekülünde herhangi bir zarar meydana getirmez. Topo-I enzimi pozitif
süpersarmal DNA'da aynı tarzda etkili değildir. Topoisomeraz
II: Bu enzim (Topo-II), ATP'den enerji sağlayarak, süpersarmal DNA'nın her iki
iplikciğinde kısa bir süre için kopmalar (açılmalar, çentikler) yaparak molekülü
istirahat haline getirir. Bakterilerde
(E. coli, vs.) bulunan ve bu sınıfta yer alan
Topo-II enzimi, DNA giraz (gyrase), daha ziyade istirahat halinde bulunan DNA
molekülünü negatif süpersarmal haline getirir. Diğer bir ifade ile DNA giraz enzimi,
süpersarmal haline gelmeyi katalize eden bir fonksiyona sahip bulunmaktadır. DNA'da
oluşan çentikler çok kısa bir süre içinde hemen kapatılarak zararlı etki yapması
önlenir. Bir molekül DNA giraz 30°C 'de her dakikada yaklaşık 100 süpersarmal
oluşturabilmektedir. Enzim 400000 molekül ağırlığında ve tetramerik bir yapıdadır
(A2B2). Hücrelerde A alt ünitesinin B'den 10 defa
daha fazla olduğu açıklanmıştır. DNA
giraz enziminin bakterilerde replikasyon çatalının ilerlemesinde çok önemli
fonksiyonu bulunmaktadır. Bu enzimin inhibisyonu, DNA'nın replikasyonunu durdurur ve
bakteride ölümlere yol açar. Bu enzimin aynı zamanda transkripsiyon ve rekombinasyonda
da işlevleri bulunmaktadır. Bazı
antibiyotikler DNA giraz'ı inhibe etmektedirler: Giraz A, nalidixic acid ve oxilinic acid
ve giraz B'de coumermycin ve novobiocin tarafından inhibe edilir. Plasmidlerde
(ColEI, pBR322, vs.), fajlarda (OX174, vs) ve viruslarda (SV40, polyoma, vs.) süpersarmal
oluşumuna rastlanılmıştır. 02.12. DNA'nın Büyüklüğü Bakterilerdeki
DNA'nın uzunluğu oldukça değişiktir. DNA'nın yaklaşık ağırlığı 0.01 pg
kadardır. (1 pico gram = 10-12 g). E. coli 'de 4x106 nukleotid çifti bulunur ve moleküler
ağırlığı da 2x109 daltondur. E. coli 'de
DNA'nın uzunluğu 1.1-1.5 mm olup bakteri boyunun 400-500 katıdır. Diğer bakterilerde
ise bu ölçüler değişik bir durum gösterir. T2 fajının DNA'sı faj boyunun 500
katı olup moleküler ağırlığı 2x106 daltondur. Küçük bakteri viruslarından olan Ø
X174 DNA fajının genomunda yaklaşık 5386 baz çifti bulunur (M.A. 1.6x106). E.
coli
'de yaklaşık 3000 kadar genin bulunduğu tahmin edilmiştir. Bunlardan ancak 1000'den
fazla genin yeri saptanabilmiştir. E.coli 'de
bir gene isabet eden baz çifti sayısı 4 x 106 / 3000 = 1333 (1400)'dür. Bir gende 1400 baz çifti
bulunduğuna göre, 1400/3 = 470 kadar kodon ve yaklaşık 470 amino asitlik bir proteini
sentezleyebilecek demektir. E.coli 'de 4x106 baz çiftinin kodları yan yana yazılsa bir
sayfaya 1000 harf (baz) konsa, yaklaşık 4000 sayfalık bir kitap veya her biri 400
sayfalık 10 cilt kitap meydana gelecek demektir. İnsanda kromozom sayısı 46 olduğu
için yaklaşık 460 cilt kitap eder. B-DNA
formunda yan yana bulunan bazlar birbirinden 0.34 nm mesafede olduğuna ve ortalama boyda
bir gende yaklaşık 1400 baz bulunduğuna göre bir genin uzunluğu 1400x0.34 = 476
nm'dir. Bir nukleotid çiftinin molekül ağırlığı yaklaşık 261 dalton olduğuna
göre, bir genin molekül ağırlığı da 261x1400 = 365400 dalton kadardır. 03.Prokaryotik Ribonukleik Asit (RNA) Mikroorganizmalarda,
DNA'dan ayrı olarak hücre içinde çok önemli fonksiyonlara sahip olan diğer bir
makromolekül de ribonukleik asit (RNA)'dır. Kompozisyonu bakımından DNA'ya çok
benzerlik göstermesine karşın fonksiyonel olarak farklı bir aktiviteye sahiptir. Bazı
araştırıcılar RNA'yı tam bir genetik materyal olarak kabul etmemekte, biri genetik
RNA (genellikle, mRNA ve RNA virusları) ve diğeri de nongenetik RNA (diğer RNA'lar)
olmak üzere iki ayrı molekül olarak düşünmektedirler. Bakteriyel
RNA'nın yapısı da başlıca 3 komponentten oluşmaktadır. 1)
Pirimidin ve pürin bazları 03.01. Pirimidin ve Pürin Bazları RNA'nın
yapısında pirimidin bazlarından Sitozin (C) ve Urasil (U: 2,4-dioksipirimidin) bulunur.
Urasil, DNA'daki timinin yerini alır. Diğer bir ifade ile RNA'da timin yoktur. Urasil,
yapı bakımından timine çok benzer. Ancak timinde 5'pozisyonunda bulunan metil grubu
(CH3) yerine urasilde hidrojen (H) bulunur.
Pürin bazları aynen DNA'da olduğu gibi 2 tanedir (Adenin ve Guanin). 03.02.Pentoz Şekeri RNA'nın
bileşiminde bulunan pentoz şekeri D-riboz (C5H10O5)
yapısındadır. 03.03. Fosfat Molekülü RNA
molekülünde DNA'daki gibi aynı H3PO4 molekülü vardır. 03.04. Nukleosidler ve Nukleotidler Nukleosidler
pirimidin bazları (sitozin, urasil) ve pürin bazlarından (adenin, guanin) biri ile
pentoz şekerinin (D-riboz) birleşmesinden meydana gelmiştir. Bazların şekerle
birleşmesi aynen DNA'da olduğu gibidir. Nukleosidler,
pirimidin nukleosid ve pürin nukleosid olarak iki grupta toplanabilirler. Nukleotidler,
nukleosidlerle fosfat molekülünün birleşmesinden oluşmuşlardır (pirimidin
nukleotidler ve pürin nukleotidler). 03.05. Polinukleotid Nukleotidlerin
fosfodiester bağlarıyla birleşmesinden polinukleotidler meydana gelirler. DNA'nın
aksine, RNA tek iplikçik halindedir. Bu nedenle de DNA'ya oranla daha zayıftır ve kolay
tahrip olur. Ancak, molekül bazı bölgelerde kendi üzerine katlanarak sağlam yapı
oluşturur. Bu birleşme adenin ile urasil ve guanin ile sitozin arasında
gerçekleştirilir. 03.06. RNA Türleri Bakterilerde
hücre içindeki fonksiyonları, molekül ağırlıkları, sedimentasyon konstantları,
birbirinden farklı 4 tür RNA bulunmaktadır. Bunlar da; 1)
Mesenger RNA (mRNA) (genetik) 03.07.Mesenger RNA (mRNA) DNA
iplikçiklerinin birinden, RNA polimeraz enziminin (DNA'ya bağımlı transkriptaz)
katalitik etkisi ile, genetik bilgiler mRNA'ya aktarılır (transkripsiyon). Yeni oluşan
mRNA, kendisinin sentezine kalıplık yapan DNA iplikçiğine anti paralel bir durum
gösterir (M.A. 500.000'in üzerindedir.) Mesenger
RNA, tek iplikçik, düz ve iki ucu açık olup yapısında A,U,C ve G bazları bulunur
(timin yoktur). E. coli 'de mRNA'nın 6000
nukleotidden fazla olduğu tahmin edilmiştir. Hücre içinde RNaz'lar tarafından kolayca
tahrip edilen mRNA'nın ömrü 2-3 dakika kadardır. mRNA'nın 5'-ucunda translasyonu
başlatma sinyalini veren 30S'lik ribosomal alt üniteye bağlanan AUG kodunu ile SD
sekansları bulunur. Bu AUG kodonu, formilize olmuş metionin tRNA'nın bağlanma yeridir.
Halbuki, mRNA'nın iç kısmında bulunan AUG kodonu ise sadece met tRNA'nın spesifik
tripletidir. Mesenger
RNA'da AUG başlama kodonundan sola doğru, Shine Dalgarno (SD) sekansları olarak bilinen
nukleotid sıraları bulunmaktadır. Bunlar mRNA'nın 30S ribosomal alt ünite üzerine
sağlamca tutunmasında rolleri fazladır. 03.08. Transfer RNA (tRNA) Protein
sentezinde önemli fonksiyona sahip olan, ribosom üzerinde sıraya giren kodonların
amino asitlerine, amino acyl senthetase ile aktive olduktan sonra, bağlanarak onları
ribosomlardaki kodonuna taşıyan ve sıraya koyan küçük bir moleküldür (75-85
nukleotid). Her amino asite özel bir veya birden fazla tRNA vardır. Amino acil sentetaz
tarafından aktive edilen amino asitler yüksek enerji ile tRNA'nın 3'-OH (CCA) ucuna
bağlanırlar. tRNA için, hücre içindeki fonksiyonuna göre 4 yapraklı bir yonca (4
kollu) benzeri morfolojisi tanımlanmaktadır. Bu kolların hepsinin ayrı işlevi
vardır. Bunlardan en önemlisi, antikodon olarak fonksiyonu olanıdır. Bu bölge,
ribosom üzerinde sırada bulunan kodonu tanıyarak bağlanır ve bu kodona tekabül eden
amino asiti de birlikte taşır. 03.09. Ribosom ve Ribosomal RNA (rRNA) Üremekte
olan hücreler de sayıları yaklaşık 15000-30000 kadar bulunan ribosomların amino
asitlerin birbirleriyle birleşmesinde ve protein haline dönüşmesinde rolü çok
büyüktür. mRNA, 30S alt ünitenin oluğuna yerleşir ve 50 S'lik ribosomla
birleştikten sonra oluşan 70 S ribosom, mRNA üzerinde kayar ve her kayışta bir kodonu
sıraya koyarak ilerler. Ribosomların
yaklaşık %60 RNA ve %40'ı da spesifik proteinlerden oluşmaktadır. E. coli
'de başlıca 3 tür rRNA bulunur (5SrRNA, 23SrRNA, 16SrRNA). Ribosomların
küçük alt ünitesi (30S), mRNA'da bulunan AUG ve Shine-Dalgarno sekanslarına
bağlanmada önemi ve etkinliği fazladır. 03.10. Primer RNA (pRNA) DNA'nın
replikasyonu sırasında parental iplikçiğin 5'®3'
yönünde olanına yeni iplikçik sentezlenirken bunun sentez yönü tersine 3'¬5'
olacağından, DNA polimeraz enzimi ters olan bu yönde sentezi kesintili olarak
sürdürür. Önce, primaz (RNA polimerase)
enzimi tarafından 50-100 nukleotidden oluşan primer RNA'lar sentezlenir. Bunlar basamak
olarak kullanılarak kısa DNA sekansları (Okazaki segmentleri) oluşturulur. DNA pol.I
enzimi yardımı ile araları doldurulur ve DNA ligaz ile de birleştirilir. pRNA'ların
görevi bittikten sonra DNA polimerase tarafından dijeste edilerek çıkarılırlar. Bu
RNA, bakterideki dnaG geni tarafından kodlanır. 04. Prokaryotik DNA ile RNA Arasındaki Başlıca Farklar 1)
DNA genellikle çift iplikciklidir. RNA ise tek iplikciklidir. Ökaryotiklerde,
mRNA molekülünün 5-ucunda 7-metilguanosin trifosfat (kep) ve 3' -ucunda da poli A
(AAAA....A) bulunur. Ancak, viruslarda da böyle durumlar bulunmaktadır. Böyle olguya, prokaryotik (bakterilerde) mRNA'da rastlanamaz. Prokaryotiklerin
mRNA'sı polisistronik (bir çok genin kodlarını taşır) olmasına karşın,
ökaryotiklerin mRNA'sı monosistroniktir (tek bir genin kodlarını taşır). 05.Nukleik Asitlerin İn Vivo
ve
İn
Vitro
Fonksiyonları 05.01. İn Vivo Fonksiyonları Nukleik
asitlerin (DNA veya RNA) hücrelerde çok önemli vital fonksiyonları bulunmaktadır.
Genetik materyallerin, canlılarda kalıtsal karakterleri taşıması ve nesillere
transferi gibi görevleri yanı sıra, hücrelerdeki bütün fizyolojik, biyokimyasal,
metabolik, morfolojik, antijenik, v.s. özelliklerin belirlenmesinde de önemli görevlere
sahiptirler. Bunlardan ayrı olarak da, genetik materyaller, enerji taşımaları,
enzimlerin kofaktörleri olmaları, kimyasal mesenger oluşturmaları, nukleik asitlerin
sentetik analoglarının kanser sağaltımında ve viral infeksiyonlarında önleme amacı
ile kullanılması gibi etkinlikleri de bulunmaktadır. Bu
son aktiviteleri hakkında aşağıda kısa ve özlü bilgiler verilmektedir. 1)
Nukleotidler kimyasal enerji bağları taşırlar: Nukleik asitlerin yapısında fosfat
moleküllerinin bulunması nedeniyle yüksek enerjiye sahip makromoleküllerdir. Bu
enerji, hücrelerde, bir çok biyokimyasal olgularda ve özellikle, biyosentezde fazlaca
kullanılırlar. Nukleotidlerde, 1, 2 ve 3 adet fosfat grubu bulunabilir (NMP, NDP ve
NTP). Bunlar içinde en önemlisi ve fazla kullanılanı 3 fosfat molekülü taşıyan
Adenosine trifosfattır (ATP). Diğerleri (UTP, GTP ve NTP) ise bazı spesifik
reaksiyonlarda gerekli olmaktadırlar. Hücre
içinde, biyosentez olaylarında fonksiyonları bulunan ve yüksek enerjiye sahip
makromoleküllerden olan nukleik asitler, yapılarında pürin ve pirimidinin yer alması
nedeniyle de DNA ve RNA'nın monomerik prekürsörlerini de oluştururlar. 2)
Nukleotidler birçok enziminin de kofaktörleridir: Hücre içinde çok önemli ve hayati
rollere sahip olan enzimlere ait kofaktörlerin yapısında da nukleotidler bulunur.
Böylece enzimlerin aktivasyonunda ve fonksiyonel bir duruma gelmelerinde nukleotidlerin
önemi oldukça fazladır. Örn, koenzim A (3' -fosfoadenin difosfat (PADP); nikotinamid
adenindinukleotid (NAD); flavin adenindinukleotid (FAD) bunlar arasındadır. 3)
Bazı nukleotidler hücrelerde sinyal iletişiminde sekonder mesenger görevi yaparlar:
Hücrelerde, nukleotidlerin diğer önemli fonksiyonları arasında bazı sinyalleri
transferde sekonder mesenjer görevi yapmaları da bulunmaktadır. Bunlardan en önemlisi,
cyclic adenosine monophosphattır (cAMP). Buna karşın guanosine monophosphatın (GMP)
etkinliği daha zayıftır. cAMP, adenilate siklase enziminin katalitik aktivasyonu sonu
ATP'den elde edilir. 4)
Sentetik nukleotid analogları klinik hekimlikte fazlaca kullanılma alanı
bulmuşlardır: Pürin ve pirimidinlerin sentetik analogları onkolojide ve viral
infeksiyonların sağaltımında, son yıllarda fazlaca yararlanılan kimyasallar
arasında yer almaktadırlar. Bu bileşikler, ya nukleik asit sentezinde önemli
fonksiyonları olan enzimlerin inhibisyonunu sağlayarak veya nukleik asitlerin yapısına
girerek, etkilerini gösterirler. Bunlar arasında, 5-fluorouracil (5-FU);
5-iodo-2-deoxyurdine; 6-mercaptopurine; 6-thioguanine 5- veya-6-azauridine;
5-veya-6-azacytidine; 8-azaguanine; allopurinol; cytarabine; azathiopurine, vs.
bulunmaktadır. 5)
Nukleotidler oksidatif fosforilasyon reaksiyonunda adaptör olarak (ATP) görev yaparlar. 6)
DNA ve RNA'nın yapısını oluşturan nukleotidlerin, genetik bilgileri taşıdıkları
ve transfer ettikleri için de önemlileri çok fazladır. 05.02. İn Vitro Fonksiyonları Nukleik
asitlerin in vitro bir çok önemli çalışmalarda da rolleri bulunmaktadır. Bunlar
kısaca şöyle sıralanabilirler. 1)
Hibridizasyon denemelerinde (türlerin sınıflandırılmalarında, genlerle ilgili
çalışmalar da, vs) [1] Kaynak: Temel Mikrobiyoloji
|
||||||||||||||||||||